Молекулярные структуры, ответственные за транспорт калия в МХ
АТФ-ингибируемые калий-транспортирующие каналы
АТФ-чувствительный калиевый канал внутренней мембраны МХ
Модуляторы митоКАТФ канала
Механизмы защиты сердца при гипоксии, опосредованные активацией митоКАТФ
Феномен прерывистой гипобарической тренировки
Выделение МХ печени крысы
Подготовка белка с м.м. 55 кДа: выделение и очистка
Исследование ДНФ-индуцированного выхода К+ из митохондрий с помощью К+-селективного электрода
Очистка антител к АТФ-зависимому белку с м.м. 55 кДа на колонке с иммобилизованным Белком А
Изучение параметров АТФ-зависимого транспорта К+ в МХ печени сердца крыс с различной резистентностью к гипоксии
Изучение структурной организации митохондриального АТФ-зависимого калиевого канала
Навигация
Изучение структурной организации митохондриального АТФ-зависимого калиевого канала
Параметры функционирования митоКАТФ у животных с различной устойчивостью к гипоксии, а также у крыс, адаптированных к кислородному голоданию
118823
знака
4
таблицы
24
изображения
5.2 Изучение структурной организации митохондриального АТФ-зависимого калиевого канала
Изучение структурной организации митоКАТФ, учитывая его существенную физиологическую роль, особенно при гипоксии, является актуальной проблемой. Как было сказано в «Обзоре литературы», японские ученые полагают, что по структуре он близок к цитоплазматическому каналу [Suzuki et al., 1997]. Однако, проведенный в лаборатории проф. Гарлида анализ действия АТ к цито-KIR на АТФ-зависимый транспорт К+ в МХ не подтвердил эти данные [Grover and Garlid, 2000]. С другой стороны, в группе проф. Марбана было высказано предположение о том, что канал образован мультикомплексом, сосоящим из 5 МХ белков, одним из которых является АВС [Ardehali et al., 2004].Следовательно, структурная организация МХ АТФ-зависимого калиевого канала до настоящего времени окончательно не выяснена.
5.2.1 Определение гомологии белка с м.м. 55 кДа методом MS-MALDI-TOF/TOF
Первым этапом изучения белка с м.м. 55 кДа было определение гомологичности его структуры последовательностям известных белков. Для этого электрофоретически чистый белок исследовали MS-MALDI-TOF/TOF (от англ. mass spectrometry-matrix assisted laser desorbtion/ionization – time of flight/time of flight) анализом.
Согласно данным, полученным MS-MALDI-TOF/TOF анализом, белок с м.м. 55 кДа на 54% гомологичен белку-предшественнику, который, исходя из результатов анализа базы данных NCBI (программное обеспечение - MASCOT (MartixScience, Москва)), является предшественником кальретикулина. Вероятно, белок с м.м. 55 кДа является конечным продуктом систем посттрансляционной модификации данного белка-предшественника. На рисунке 14 представлена аминокислотная последовательность белка-предшественника. Последовательности данного белка, перекрывающиеся с последовательностями 55 кДа белка, выделены серым цветом.
1 MLLSVPLLLG LLGLAAADPA IYFKEQFLDG DAWTNRWVES KHKSDFGKFV
51 LSSGKFYGDQ EKDKGLQTSQ DARFYALSAR FEPFSNKGQT LVVQFTVKHE
101 QNIDCGGGYV KLFPGGLDQK DMHGDSEYNI MFGPDICGPG TKKVHVIFNY
151 KGKNVLINKD IRCKDDEFTH LYTLIVRPDN TYEVKIDNSQ VESGSLEDDW
201 DFLPPKKIKD PDAAKPEDWD ERAKIDDPTD SKPEDWDKPE HIPDPDAKKP
251 EDWDEEMDGE WEPPVIQNPE YKGEWKPRQI DNPDYKGTWI HPEIDNPEYS
301 PDANIYAYDS FAVLGLDLWQ VKSGTIFDNF LITNDEAYAE EFGNETWGVT
351 KAAEKQMKDK QDEEQRLKEE EEDKKRKEEE EAEDKEDEDD RDEDEDEEDE
401 KEEDEEDATG QAKDEL
Рис.12. Аминокислотная последовательность прекурсорного белка. Участки структуры белка-предшественника, совпадающие с последовательностями, имеющимися в белке с м.м. 55 кДа, выделены серым цветом
При рассмотрении структуры типичного кальретикулина, выделенного из печени крысы, процент перекрывания его аминокислотных последовательностей с известными последовательностями 55 кДа белка также оказался достаточно большим (Рис.13).
1 mllsvplllg llglaaadpa iyfkeqfldg dawtnrwves khksdfgkfv
51 lssgkfygdq ekdkglqtsq darfyalsar fepfsnkgqt lvvqftvkhe
101 qnidcgggyv klfpggldqk dmhgdseyni mfgpdicgpg tkkvhvifny
151 kgknvlinkd irckddefth lytlivrpdn tyevkidnsq vesgsleddw
201 dflppkkikd pdaakpedwd erakiddptd skpedwdkpe hipdpdakkp
251 edwdeemdge weppviqnpe ykgewkprqi dnpdykgtwi hpeidnpeys
301 pdaniyayds favlgldlwq vksgtifdnf litndeayae efgnetwgvt
351 kaaekqmkdk qdeeqrlkee eedkkrkeee eaedkededd rdededeede
401 keedeedatg qakdel
Рисунок 13. Аминокислотная последовательность типичного кальретикулина (21-332 – кальретикулин, 1-17 – сигнальная последовательность, удаляемая протеолизом при созревании белка). Участки последовательности, общие с белком с м.м. 55 кДа, выделены серым цветом
Классический кальретикулин входит в состав семейства высококонсервативных белков с м.м. около 55 кДа, локализующихся как в саркоплазматическом, так и в эндоплазматическом ретикулуме клеток печени, скелетной и гладкой мускулатуры, сердечной мышцы [Fliegel et al., 1989]. Белки этого семейства связывают ионы кальция, цинка, других металлов, сахара, нуклеотидфосфаты, в частности, УДФ. Они также выступают в роли шаперонных белков в процессе фолдинга, участвуют в системе белковой машинерии, регуляции апоптоза, мейоза, экспорта белков из ядра клетки. Известны гомологи гена кальретикулина у мыши, крысы и человека. У крысы гомологичный ген кальретикулина локализован в 19 хромосоме. При этом ранее в нашей лаборатории было показано, что при окрашивании белков в геле после ДДС-ПААГ электрофореза по Шиффу, исследуемый белок с м.м. 55 кДа давал положительную реакцию, что говорит о способности его связывать сахара [неопубликованные данные]. Кроме того, определялось также и сродство белка-канала к кальцию с использованием 45Ca [неопубликованые данные]. Было показано, что исследуемый белок с м.м. 55 кДа проявляет достаточно большое сродство к этому иону. Эти результаты согласуются с данными по взаимодействию кальретикулина с сахарами и кальцием.
Причем, необходимо заметить, что большая часть перекрывающейся последовательности приходится на Р-домен кальрегулина, способного связывать нуклеотиды. Также, в перекрывающуюся последовательность включены высоконсервативные остатки триптофана, характерные для кальрегулинов в данных сайтах первичной последовательности, и стерически идентично расположенные в первичной аминокислотной последовательности кальрегулина консервативные антипараллельные бета-листы, формирующие Р-домен.
Следует особенно отметить тот факт, что в составе прекурсорного белка и кальретикулинов различных типов имеется характерная гидрофобная сигнальная последовательность из 17 аминокислотных остатков, MLLSVPLLLGLLGLAAA (1-17), посттрансляционно удаляемая протеолитическим расщеплением [Murthy et al., 1990] (Рис.16). Эта последовательность отсутствует в изучаемом нами белке с м.м. 55 кДа. Интересно также то, что N-концевая последовательность с 17 по 27 аминокислотный остаток DPAIYFKE кальретикулина совпадает с последовательностью N-концевого участка 55 кДа белка, структура которого была определена ранее в нашей лаборатории реакцией химической деградации по Эдману [неопубликованные данные] (Рис.16).
Кроме того, С-конец прекурсорного белка, как и всех кальретикулинов, содержит сигнальную последовательность KDEL, определяющую локализацию этих белков в ретикулюме [Schweizer et al., 1993], в то время как у изучаемого 55 кДа белка такой последовательности нет (Рис.15, 16), что, вероятно, свидетельствует о том, что белок с м.м. 55 кДа локализуется не в ретикулюме.
1 MLLSVPLLLG LLGLAAADPA IYFKEQFLDG DAWTNRWVES KHKSDFGKFV
51 LSSGKFYGDQ EKDKGLQTSQ DARFYALSAR FEPFSNKGQT LVVQFTVKHE
101 QNIDCGGGYV KLFPGGLDQK DMHGDSEYNI MFGPDICGPG TKKVHVIFNY
151 KGKNVLINKD IRCKDDEFTH LYTLIVRPDN TYEVKIDNSQ VESGSLEDDW
201 DFLPPKKIKD PDAAKPEDWD ERAKIDDPTD SKPEDWDKPE HIPDPDAKKP
251 EDWDEEMDGE WEPPVIQNPE YKGEWKPRQI DNPDYKGTWI HPEIDNPEYS
301 PDANIYAYDS FAVLGLDLWQ VKSGTIFDNF LITNDEAYAE EFGNETWGVT
351 KAAEKQMKDK QDEEQRLKEE EEDKKRKEEE EAEDKEDEDD RDEDEDEEDE
401 KEEDEEDATG QAKDEL
Рисунок 14. Последовательность прекурсорного белка. Сигнальная последовательность, характерная для кальретикулинов, выделена серым цветом, N-концевая последовательность, гомологичная таковой у исследуемого белка-канала с м.м. 55 кДа – жирным шрифтом, сигнальная последовательность KDEL, остутствующая в изучаемом белке – серым цветом и жирным шрифтом
Следует также отметить, что кальретикулины содержат ряд высоко консервативных последовательностей [Fliegel et al., 1989]. Согласно результатам MS-MALDI-TOF/TOF анализа, ряд гомологичных с белком предшественником участков цепи белка с м.м. 55 кДа являются высококонсервативными для белков семейства кальретикулинов.
Возможно, исследуемый нами белок, гомологичный кальрегулину, проходит альтернативные стадии пострансляционных модификаций и созревания при биосинтезе. Это, вероятно, позволяет ему встраиваться во внутреннюю мембрану митохондрий беспрепятственно и без значительных потерь энтропии и внутренней энергии при смене гидрофильной среды на гидрофобную. Не исключено также дополнительное стерическое и физико-химическое влияние на химические свойства и конформационную норму со стороны компонентов мембраны митохондрий. Учитывая все вышесказанное, на данном этапе исследования структуры митоКАТФ, можно утверждать лишь, что белок с м.м. 55 кДа является белком, обладающим высокой степенью структурной и функциональной гомологии с кальрегулином.
Учитывая все вышесказанное, на данном этапе исследования структуры митоКАТФ, можно утверждать лишь, что белок с м.м. 55 кДа является белком, обладающим высокой степенью структурной и функциональной гомологии с кальрегулином.
5.3 Ингибиторный анализ активности митоКАТФ канала с использованием антител, полученных на белок с м.м. 55 кДа
В нашей лаборатории были получены косвенные доказательства гетеромультимерного строения этого канала, а также данные о том, что белок с м.м. 55 кДа является канальной субъединицей митоКАТФ [Григорьев, 1999; Негода, 2004; Mironova et al., 2004]. В то же время, результаты исследования гомологичности структуры белка-канала с м.м. 55 кДа, выделенного из МХ печени крысы, показали высокий процент гомологии исследуемого белка с кальрегулином. Таким образом, возникла необходимость доказательства принадлежности белка с м.м. 55 кДа к системе митохондриального АТФ-зависимого транспорта калия.
Для прямого доказательства принадлежности каналообразующего белка с м.м 55 кДа к митохондриальной системе АТФ-ингибируемого транспорта К+, в работе на этот белок были получены поликлональные антитела (АТ) и проведен анализ их влияния на АТФ-зависимый транспорт калия в интактных МХ.
Предварительные данные по влиянию АТ к 55 кДа белку на транспорт К+ в МХ были получены в лаборатории ранее [Скарга и др., 1986]. Однако в о время, белок не был идентифицирован как АТФ-зависимый К+ канал и методы его очистки были несовершенны. Кроме того, не все использовавшиеся ранее модели отражали работу АТФ-зависимого митоходнриального калиевого кнала. Не было также проведено сравненительное исследование по влиянию АТ к 55 кДа белку на другие функции МХ.
5.3.1 Определение степени чистоты белка, используемого для иммунизацииДля получения поликлональных специфических антител на белок-канал необходимо было получить гомогенный белок с м.м. 55 кДа. Этот белок выделяли из МХ печени крысы методом водно-этанольной экстракции [Миронова и др., 1981; 1996(I)]. Дальнейшую очистку белка проводили методом ионообменной хроматографии с использованием ДЭАЭ-целлюлозы (п.п. 2 «Материалов и методов»). Каналообразующий белок элюировался с колонки 250 мМ KCl. Чистота данной фракции определялась ДДС-ПААГ электрофорезом [Laemmli, 1979] (Рис.5). Для обнаружения активной фракции, все фракции, элюированные с колонки ДЭАЭ-целлюлозы тестировали при встраивании в БЛМ (см. «Материалы и методы»). Далее активную фракцию (250 мМ KCl) диализовали против 5 мМ Трис буфера (pH 7.2) с добавлением 0.1% b-меркаптоэтанола в течение ночи при 4°С. Диализат повторно наносили на колонку ДЭАЭ-целлюлозы, аналогичную использовавшейся в первом случае. Фракции элюировали ступенчатым градиентом KCl. Все фракции тестировались на активность при встраивании в БЛМ. АТФ-ингибируемые К+-селективные каналы формировались, как правило, при встраивании белка, элюированного 250 мМ KCl.
Дополнительная очистка проводилась методом препаративного нативного электрофореза (п.п. 5.1.1. «Материалов и методов») в 10%-ом ПААГ. На конечной стадии очистки, исследуемый белок элюировали из геля, концентрировали методом обратного диализа на ПЭГ и проверяли на ДДС-ПААГ электрофорезе (Рис.17). Электрофоретическая подвижность белка соответствовала массе ~55 кДа.
|
|
|
|
|
|

Рис. 15. Результат ДДС-ПААГ электрофореза фракции, полученной после нативного электрофореза и содержащей белок с м.м. 55 кДа. 1 – стандарт молекулярных масс, 2 – фракция митоКАТФ канала после нативного электрофореза
Полученным электрофоретически гомогенным белком с м.м. 55 кДа модифицировали искусственные бислойные мембраны. Было показано, что данный белок обладает селективной К+-транспортирующей активностью и ингибируется физиологическими концентрациями АТФ (Рис.18).
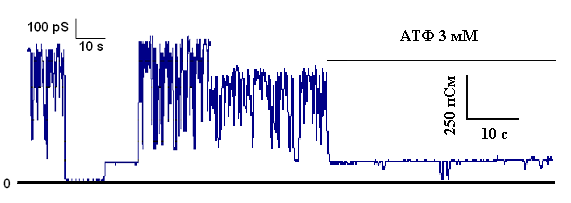
Рисунок 16. К+-транспортирующая АТФ-ингибируемая активность белка с м.м. 55 кДа, реконструированного в искусственную мембрану. Среда инкубации содержала: 20 мМ Трис, 100 мМ KCl, рН 7.2
Электрофоретически гомогенный АТФ-ингибируемый К+-транспортирующий белок с м.м. 55 кДа и замораживали и накапливали для иммунизации. При выделении вышеописанным методом из 100 г печени крыс получали 30-70 мкг очищенного белка. Иммунизацию кроликов проводили при накоплении 0.2-0.4 мг белка.
5.4 Иммунизация и определение титра полученных антителПеред началом иммунизации отбирали 30-40 мл крови каждого животного для приготовления нормальной контрольной сыворотки. Для получения антител к АТФ-ингибируемому К+-транспортирующему белку с м.м. 55 кДа кроликов иммунизировали электрофоретически чистым белком. С целью устранения индивидуальных различий в получаемых анителах иммунизировали двух кроликов массой 2.6-2.8 кг.
Схема эксперимента была подобрана в соответствии с требованиями Декларации совета Европейского Союза 86/609/EEC и представлена в таблице 8. Инъекции делали подкожно в область лопаток в 3 подхода. Интервал между инъекциями 10-15 дней.
В ходе иммунизации кроликов по представленной выше схеме были получены антитела к белку с м.м. 55 кДа, выделенному из МХ печени крысы. Определение титра антител, полученных на белок-канал с м.м. 55 кДа проводили методом непрямого дот-анализа (см. «Материалы и методы»). Средние значения титра антител указаны в таблице 8.
Таблица 2. Схема иммунизации кроликов АТФ-зависимым К+-транспортирующим белком м.м. 55 кДа.
| Номер животного | Кролик №1 | Кролик №2 | ||||
| Номер иммунизации | 1 | 2 | 3 | 1 | 2 | 3 |
| Количество белка, мкг | 114 | 70 | 30 | 100 | 114 | 38 |
| Суммарное количество белка, мкг | 214 | 252 | ||||
| Титр после трех иммунизаций | 1:3200 | 1:1600 | ||||
Специфичность антител на белок с м.м. 55 кДа определяли методом Вестерн-блот. Для этого выделенный из МХ печени крысы и очищенный АТФ-чувствительный К+-транспортирующий белок с м.м. 55 кДа наносили на ДДС-ПААГ электрофорез с 10%-ым ПААГ гелем с нагрузкой на полосу 10 мкг. На другую полосу наносили 10 мкг тотальной белковой фракции МХ. Порядок нанесения образцов дублировался на том же геле. После электрофоретического разделения белков половина геля, содержащая оба образца и стандарт молекулярных масс, окрашивалась серебрением [Shevchenko et al., 1996]. Вторая половина использовалась для переноса на нитроцеллюлозную мембрану с последующей обработкой полученными антителами (п.п. 5.2.2. «Материалов и методов»). При этом исследуемую антисыворотку разводили 1:300, вторичные антитела – 1:500.
|
|
|
|


Рисунок 17. А. ДСН-электрофорез фракций
1 – очищенный АТФ-чувствительный К+-транспортирующий белок с м.м. 55 кДа, 2 – молекулярные стандарты массы белков, 3 – суммарный белок МХ. Б. Вестерн-блот анализ: 1 – очищенный АТФ-чувствительный К+-транспортирующий белок с м.м. 55 кДа, 2 – суммарный белок МХ.
Показано, что полученные в работе антитела специфически связываются только с белком с м.м. 55 кДа, и не вязываются ни с одним из массы белков, находящихся в МХ (Рис.19). Таким образом, в работе были получены специфические поликлональные антитела на белок с м.м. 55 кДа.
Этим же методом было установлено, что поликлональные антитела на исследуемый белок-канал, полученный из МХ печени крысы, не взаимодействует с той же концентрацией аналогичного белка с м.м. 55 кДа, выделенного тем же методом из МХ сердца крысы. Данный факт указывает на то, что исследуемый белок с м.м. 55 кДа тканеспецифичен.
5.4.2 Выделение иммуноглобулинов G (IgG) из антисыворотки и проведение ингибиторного анализаПолученные антитела к АТФ-чувствительному К+-транспортирующему белку с м.м. 55 кДа, выделенному из МХ печени крысы, использовали в качестве ингибитора АТФ-зависимого транспорта ионов калия в МХ печени крысы. Для этого, полученные очищенные и концентрированные антитела (иммуноглобулины G (IgG)) кроликов (см. «Материалы и методы»). Контролем служили очищенные IgG сыворотки крови Интактных (неиммунизированных) кроликов. Кроме того, в качестве контроля использовались антитела к белку с м.м. 55 кДа, подвергнутые кипячению в течение 5 минут. Ранее было показано, что такая процедура ведет к потере белком активности. IgG контрольных животных получали тем же способом, что и IgG рабочей сыворотки. В ходе выделения и очистки IgG титр антител и сродство к белку-каналу с м.м. 55 кДа существенно не изменяется. Степень чистоты выделенных из сыворотки IgG определяли методом ДДС-ПААГ электрофореза [Laemmli, 1979] (см. «Материалы и методы»).
Очищенные фракции IgG, выделенные из иммунизированных и интактных животных, использовали для ингибирования АТФ-зависимого выхода ионов К+ из МХ в присутствии ДНФ, отражающего работу митоКАТФ, и энергозависимого входа ионов К+ в МХ. Все эксперименты проводились при термостатировании (26°С) и постоянном перемешивании. После 1.5-2 минут преинкубации антител с митохондриями транспорт ионов К+ индуцировали ДНФ (при исследовании выхода К+ из МХ К+-селективным электродом) или субстратом дыхания (при определении энергозависимого входа К+ в МХ). Для повышения эффективности взаимодействия IgG с внутренней мембраной МХ создавались гипотонические условия. На рисунке 6 представлена концентрационная зависимость степени ингибирования выхода ионов К+ из МХ печени крысы в присутствии разобщителя (ДНФ). Ингибирующий эффект антител наблюдается только в случае добавления в среду инкубации МХ очищенных интактных IgG к АТФ-зависимому К+-транспортирующему белку с м.м. 55 кДа, выделенному из печени крысы. Степень ингибирования зависела от концентрации IgG и времени преинкубации.
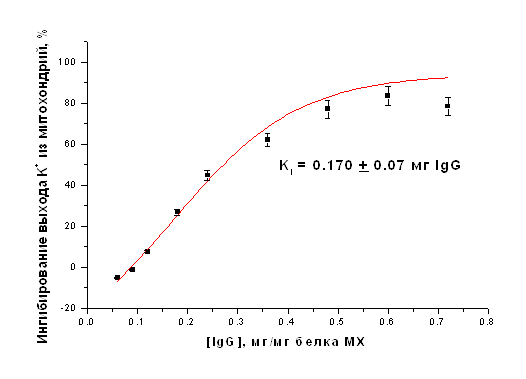
Рисунок 18. Ингибирование АТФ-зависимого ДНФ-индуцированного выхода К+ из МХ печени крысы антителами на АТФ-чувствительный К+ белок-канал с м.м. 55 кДа. Кi = 0.170+0.07 мг IgG/мг белка МХ
Максимальное ингибирование, наблюдавшееся в данных экспериментах составляло 83%. Константа полумаксимального ингибирования (Кi) составила 0.170+0.07 мг IgG/мг белка МХ (Рис. 20).
Антитела к белку с м.м. 55 кДа, выделенному из МХ печени крысы, ингибируют также энергозависимый вход ионов К+ (Рис. 21).
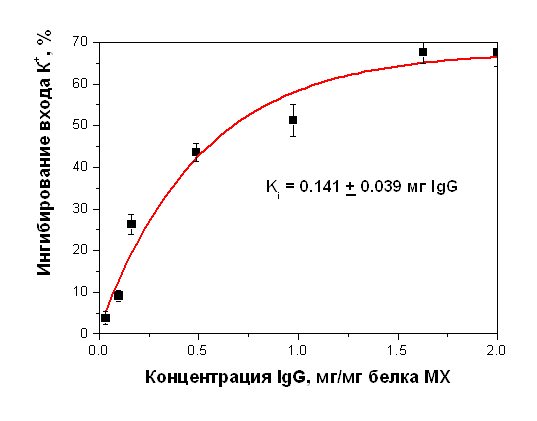
Рисунок 19. Ингибирование энергозависимого входа ионов К+ в МХ печени крысы антителами на белок с м.м. 55 кДа
Следует отметить, что IgG контрольной сыворотки и IgG, инактивированные кипячением, не влияли ни на ДНФ-индуцированный выход калия из МХ, ни на энергозависимый вход ионов (Таблица 9). Кроме того, антитела, полученные на АТФ-зависимый К+-транспортирующий белок с м.м. 55 кДа, выделенный из МХ печени крысы, не блокировали АТФ-чувствительный К+ транспорт в МХ сердца крысы, при использовании обоих методов исследования (Таблица 9).
Таблица 3. Процент максимального ингибирования АТФ-зависимого К+ транспорта в МХ печени и сердца крыс антителами, полученными на белок с м.м. 55 кДа, выделенный из МХ печени крысы.
| Образец | % максимального ингибирования АТФ-зависимого входа К+ в МХ, измеренного методом спектрофотометрии | % максимального ингибирования ДНФ-индуцированного выхода К+ из МХ, измеренного с помощью К+-селективного электрода |
| МХ печени | 68 | 80 |
| МХ сердца | 1.5 | 2 |
| МХ печени + АТ преимунной сыворотки | 0 | 0 |
| МХ сердца + АТ преимунной сыворотки | 0 | 0 |
| МХ печени + АТ, инактивированные 5-тиминутным кипячением | 1 | 1.5 |
В работе также определяли влияние полученных антител на параметры дыхания МХ. Ни контрольные IgG, ни IgG, специфические к белку-каналу с м.м. 55 кДа (Рис. 22), ни инактивированные IgG, не оказывали заметного влияния на дыхание МХ в присутствии субстрата и АДФ.
Отсутствие подобного влияния связано, по-видимому, с тем, что в интактных МХ, в которых не индуцировали транспорт К+, роль данного иона в процессе дыхания незначительна.
Таким образом, в работе получены специфические поликлональные антитела на выделенный из МХ печени крысы белок с м.м. 55 кДа, формирующий при встраивании в БЛМ АТФ-ингибируемые К+-селективные каналы. Данные антитела блокируют АТФ-зависимый вход К+ в нативные МХ и ДНФ-индуцированный, ингибируемый АТФ, выход этого иона из МХ. Следует отметить, что исследуемые антитела не влияют на другие параметры функционирования МХ, в том числе, на дыхание. Полученные результаты доказывают, что белок с м.м. 55 кДа действительно участвует в формировании АТФ-зависимого калиевого канала внутренней мембраны МХ и, по всей видимости, является его канальной субъединицей.
5.5 Электронная микроскопия МитоКАТФ каналаДля более точного доказательства локализации белка с м.м. 57 кДа во внутренней мембране митохондрий было проведено электронно-микроскопическое исследование срезов тканей печени и сердца с использованием АТ на белок с м.м. 55 кДа. Идентификации эндогенного белка-канала и его связывание с АТ проводились с помощью вторичных антител, меченых коллоидным золотом (диаметр гранул – 10 нм). Как видно из рисунка 21, такого рода гранулы золота локализуются во внутренней мембране митохондрий как печени (Рис.21а), так и сердца (Рис.21б), причем в сердце их больше, что соответствуют данным о большей плотности митоКАТФ каналов в митохондриях сердца. Локализуются эти каналы ближе к месту контакта внутренней и внешней мембраны митохондрий, что особенно проявляется с митохондриях сердца (Рис.21б).
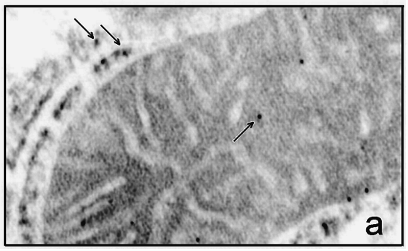
Рис.21а. Электронная микроскопия среза гепатоцита крысы. а - Срезы инкубированы с антителами к митохондриальному К+-транспортирующему белку. Черные гранулы (указаны стрелкой) -сайты локализации К+-транспортирующего белка с м.м. ~ 55 кДа в митохондриях.
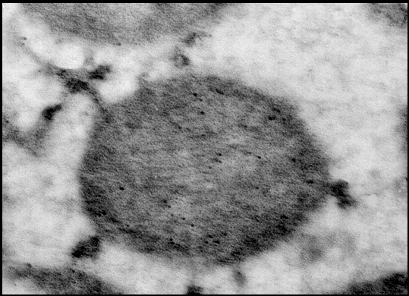
Рис.21 б. Митохондрии и саркоплазматический ретикулум (СР) на срезе кардиомицитов крысы. Срезы инкубированы с антителами к митохондриальному К+-транспортирующему белку. Черные гранулы- сайты локализации К+-транспортирующего белка с м.м. ~ 57 кДа в митохондриях, а также сайты связывания антител с СР.
Такие же гранулы (Рис.21А) обнаружены и в ретикулюме, особенно в месте его слияния с мембраной митохондрий, что подтверждает выявленную нами методом MS-MALDI-TOF/TOF – анализа общность в структуре изучаемого белка с микросомах кальретикулином (м.м. 55 кДа).
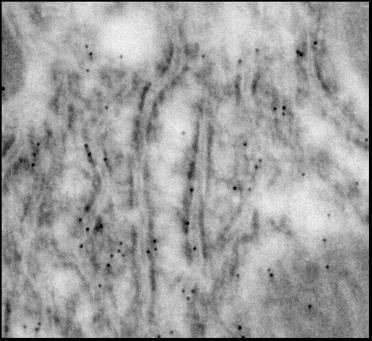
Рис.22. Электронная микроскопия эндоплазматический ретикулум (ЭР) на срезе гепатоцитов крысы. Срезы инкубированы с антителами к митохондриальному К+-транспортирующему белку.
В контрольных экспериментах, где были использованы только вторичные АТ, гранулы коллоидного золота не были выявлены. Использование антител к канальной субъединице цитоплазматического КАТФ канала – KIR6.2 не обнаружило наличия этого белка-канала в митохондриях (Рис.23, 24).
Рис.23. Электронная микроскопия среза гепатоцита крысы. Контроль (Первичные антитела заменены буфером).
Рис.24. Митохондрии и саркоплазматический ретикулум (СР) на срезе кардиомицитов крысы. б - Контроль (Первичные антитела заменены буфером)
Т.о., полученные результаты показали отсутствие общности в структуре KIR6.2 и изучаемого нами митохондриального белка-канала.
Приведенные в настоящей работе данные позволяют подтвердить, высказанное нами ранее предположение, что белок с м.м. 55 кДа относится к митохондриальной системе АТФ-зависимого транспорта калия и вместе с известными в литературе глибенкламид-связывающими белками митохондрий входит, вероятно, в состав митохондриального АТФ-зависимого калиевого канала, являясь его канальной субъединицей (митоKIR).
ЗАКЛЮЧЕНИЕ
Настоящая работа посвящена изучению роли митохондриального АТФ-чувствительного калиевого канала (митоКАТФ) в защите сердца от ишемии, формировании адаптации животных к гипоксии, а также исследованию структурной организации данного канала.
В представленной работе также была изучена роль митоКАТФ канала в формировании адаптации животных к кислородному голоданию. Для этого использовали крыс, разделенных на две группы (низкоустойчивые и высокоустойчивые) по способности выдерживать подъем на высоту в 11500 м. Кроме того, в работе низкоустойчивые животные, были адаптированы к недостатку кислорода прерывистой нормобарической гипоксической тренировкой.
Показано, что у высокоустойчивых животных митоКАТФ канал работает эффективнее, а показатели сопряжения дыхательной цепи были выше, чем у низкоустойчивых. Адаптация низкоустойчивых животных к гипоксии сопровождается сопряжением дыхательной цепи и активацией митоКАТФ канала. В то же время, поскольку набухания, которое, как предполагается, должно следовать за активацией митоКАТФ, не наблюдается, так как количество калия в митохондриях, по нашим данным, не увеличивается, а даже снижается, вероятнее всего, активируется также и система выхода калия из митохондрий (система К+/Н+-обменника).
Задачей настоящей работы было также выяснение структурной гомологии белка с м.м. 55 кДа, формирующего канальную субъединицу митоКАТФ канала [Mironova et al., 2004] аминокислотным последовательностям известных белков. Для этого проводился MS-MALDI-TOF/TOF анализ, с последующей обработкой результатов. Было показано, что белок с м.м. 55 кДа обладает высокой (54%) степенью структурной и функциональной гомологии с типичным представителем семейства кальрегулинов.
Полученный результат вызвал необходимость дополнительного исследования принадлежности исследуемого белка-канала к семейству митохондриальных белков. Для этого на 55 кДа белок, выделенный из митохондрий печени крысы и формирующий при встраивании в искусственные мембраны АТФ-чувствительный калиевый канал, были получены специфические поликлональные антитела. Вестерн-блот анализ с полученными антителами позволил выявить тканеспецифичность изучаемого белка-канала. Был также проведен ингибиторный анализ АТФ-зависимого транспорта калия в митохондриях, результаты которого показали, что АТ на белок из митохондрий печени крыс ингибировали этот транспорт в митохондриях печени, но не сердца крыс. При этом они не влияли на показатели сопряженного дыхания митохондрий печени крыс. Следовательно, белок с м.м. 55 кДа действительно относится к системе АТФ-чувствительного входа К+ в митохондрии. Для более точного доказательства локализации белка с м.м. 57 кДа во внутренней мембране митохондрий было проведено электронно-микроскопическое исследование срезов тканей печени и сердца с использованием АТ на белок с м.м. 55 кДа. Которое так же подтверждает высказанное ранее предположение.
1. Формирование устойчивости животных к недостатку кислорода, а также адаптация нормобарической гипоксической тренировкой низкоустойчивых животных сопровождается сопряжением дыхательной цепи, активацией митоКАТФ канала и К+/Н+ - обменника.
2. Показано, что белок с м.м. 55 кДа обладает высокой степенью структурной и функциональной гомологии с типичным представителем семейства кальрегулинов.
3. В работе получены специфические поликлональные антитела на белок с м.м. 55 кДа, формирующий при встраивании в искусственную мембрану АТФ-ингибируемые К+ каналы. Ингибиторный анализ АТФ-чувствительного калиевого транспорта в интактных МХ с использованием полученных антител показал, что белок с м.м. 55 кДа относится к системе АТФ-зависимого входа К+ митохондрий.
4. Электронно-микроскопическое исследование срезов тканей печени и сердца крысы после их инкубации с АТ на белок с м.м. 55 кДа, выделенный из внутренней мембраны митохондрий печени показало его принадлежность к системе митохондриального транспорта калия.
ЛИТЕРАТУРА
гипоксия белок калий антитело
1. Баграмян К., Трчунян А. Особенности структуры и функционирования формиат-водород-лиазы-фермента смешанного брожения у Escherichia coli. // Биохимия, 2003, т. 68, № 11, с. 1445-1458.
2. Баграмян К.А. Электрохимическое исследование протон-транслоцирующей функции гидрогеназы 3. // Биофизика, 2002, т. 47, № 5. с.847-851.
3. Баранова О.В., Скарга Ю.Ю., Негода А.Е., Миронова Г.Д. Ингибтрование адениновыми нуклеотидами ДНФ-индуцированного транспорта калия в митохондриях. // Биохимия, 2000, т. 65, № 2, с. 262-267.
4. Брустовецкий Н.Н., Данилова Л.С., Маевский Е.И., Колаева С.Г. Изменения реакций окислительного фосфорилирования в митохондриях печени крыс и суслтков при адаптации к холоду и в состоянии зимней спячки. // Эволюционные аспекты гипобиоза и зимней спячки. 1986, с. 69-72
5. Долгов В.В., Райскина М.Е., Антонов В.Ф. Действие адреналина на содержание К+ в митохондриях сердца собаки и зависимость транспорта К+ от дыхания и окислительного фосфорилирования. // Биофизика, 1974, т. 19, № 6, с 1025-1029.
6. Зинченко В.П., Кудзина Л.Ю., Евтодиенко Ю.В., Ким Ю.В. Характеристика К+- транспортирующей системы митохондрий при интенсивной мышечной нагрузке. // Биохимия, 1982, т. 47, № 11, с. 1839-1843.
7. Иванов К.П. Современная теория терморегуляции и зимняя спячка. // Эволюционные аспекты гипобиоза и зимней спячки. 1986, с. 49-54.
8. Киракосян Г., Баграмян К., Трчунян А. Окислительно-восствновительные процессы и образование молекулярного водорода бактериями Escherichia coli в гиперосмотической среде. // Биофизика, 2001, т. 46, № 2, с. 245-250.
9. Кондрашова М.Н., Ахмеров Р.Н., Григоренко Е.В., Федотчева Н.И., Миронова Г.Д. Торможение окисления янтарной кислоты как причина снижения теплопродукции при спячке. // Эволюционные аспекты гипобиоза и зимней спячки. 1986, с. 55-60.
10. Кудзина Л.Ю., Юрков И.С., Полтева Н.А., Евтодиенко Ю.В., Кондрашова М.Н. Влияние редокс-состояния дыхательной цепи на проницаемость мембраны митохондрий для ионов калия. // Биохимия. 1981, т.46, с 1807-1814.
11. Ленский А.С., Введение в бионеорганическую и биофизическую химию. М.: Высш. Шк., 1985, 152 с.
12. Мартиросов С.М., Трчунян А.А. Взаимодействие систем транспорта Н+ и К+ у анаэробно и аэробно выщенных E. сoli. // Биофизика, 1986, т. ХХХI, № 3, с. 464-467.
13. Мартиросов С.М., Трчунян А.А. Поглощение К+ у E. Сoli, выращенных в аэробных условиях. // Биофизика, 1986, т. ХХХI, № 4, с. 626-630.
14. Маршанский В.Н., Новгородов С.А., Ягужинский Л.С. Влияние специфических ингибиторов ферментов дыхательной цепи и АТФ-синтетазы на транспорт ионов в митохондриях, индуцированный неферментативными перекисными реакциями. // Биофизика, 1983, т.28, №5, с. 830-834.
15. Маршанский В.Н., Ягужинский Л.С. Влияние субстратов АТФ-синтетазы на индукцию процесса перекисного окисления липидов в митохондриях. //. Биол. мембраны, 1985, т.2, № 11, с. 1081-1086.
16. Миронова Г.Д., Григорьев С.М. Скарга Ю.Ю., Негода А.Е., Коломыткин О.В. // . АТФ-зависимый калиевый канал митохондрий печени крысы. ІІ. Ингибиторный анализ, кластеризация канала. // Биологические мембраны, 1996 б, т. 13, № 5, с. 537-544.
17. Миронова Г.Д., Маслова Г.М., Федотчева Н.И., Миронов Г.П. Участие митохондриальных систем транспорта в термогенезе теплокровных животных. // В сб. : Эволюционные аспекты гипобиоза и зимней спячки. Л.: Наука, 1986, с.64-68.
18. Миронова Г.Д., Проневич Л.А., Федотчева Н.И.,Сирота Т.В., Трофименко Н.В., Миронов Г.П. Системы транспорта катионов в митохондриях. // Митохондриальные процессы во временной организации жизнедеятельности. Пущино, 1978, с. 451-457.
19. Миронова Г.Д., Скарга Ю.Ю., Григорьев С.М., Яров-Яровой В.М., Александров А.В., Коломыткин О.В. АТФ-зависимый калиевый канал митохондрий печени крысы. І. Выделение, очистка и реконструкция канала в БЛМ. // Биологические мембраны, 1996 а, т. 13, № 4, с. 396-403.
20. Миронова Г.Д., Федотчева Н.И., Макаров П.Р., Проневич Л.А., Миронов Г.П. Белок из митохондрий сердца быка, индуцирующий канальную калиевую проводимость бислойных липидных мембран. // Биофизика, 1981, т. 26, с. 451-457.
21. Миронова Г.Д., Федотчева Н.И., Скарга Ю.Ю., Кондрашова М.Н. Транспорт калия и дыхание митохондрий при выходе суслика из состояния зимней спячки. // Механизмы зимней спячки. Пущино, 1987, с. 39-47.
22. Миронова Г.Д., Федотчева Н.И., Скарга Ю.Ю., Копецки Я., Хоуштек И. Сравнительный анализ термогенных систем митохондрий печени и бурой жировой ткани. // Механизмы природных гипометаболических состояний. Пущино, 1991, с. 34-43.
23. Мнацаканян Н., Захарян Э., Баграмян К., Трчунян А. Дитиол-дисульфидные переходы в мембранных транспортных белках у Escherichia coli. // Биологические мембраны, 2002, т. 19, № 2, с. 183-192.
24. Скарга Ю.Ю., Долгачева Л.П., Федотчева Н.И., Миронова Г.Д. Влияние антител к митохондриальному К+- транспортирующему белку на транспорт К+ в митохондриях печени крысы. // Укр. биохим. Журн., 1987, т. 59, № 6, с. 54-59.
25. Смирнова В.Г., Красных Т.А., Октябрьский О.Н. Роль глутатиона при ответе Escherichia coli на осмотический шок. // Биохимия, 2001, т. 66, № 9, с. 1195-1201.
26. Тер-Никогосян В.А., Трчунян А.А., Мартиросов С.М. Характер поглощения К+ у анаэробно выращенных S. Typhimurium. // Биофизика, 1986, т. ХХХI, № 5, с. 825-828.
27. Трчунян А.А., Дургарьян С.С., Оганджанян Е.С., Тер-Никогосян В.А., Варданян А.Г., Оганесян М.И., Петросян Л.С., Ванян П.А., Карагулян Э.А., Мартиросов С.М. Исследование способности анаэробно выращенных бактерий обменивать 2Н+ клетки на К+ среды и поддерживать высокое распределение К+ между клеткой и средой. // Биологические науки, 1986, №12, с. 82-88.
28. Трчунян А., Оганджанян Е.., Саркисян Э., Гонян С., Оганесян А., Оганесян С. Мембранотропные эффекты электромагнитного излучения крайне высоких частот на Escherichia coli. // Биофизика, 2001, т.46, № 1, с.69-76.
29. Федотчева Н.И. Влияние ГТФ на содержание ионов К+ и окисление субстратов в митохондриях бурого жира. // Механизмы природных гипометаболических состояний. Пущино, 1991, с. 43-49.
30. Федотчева Н.И., Мирзабеков Т.А., Миронов Г.П., Миронова Г.Д. Изменения транспорта К+ в митохондриях печени сусликов при зимней спячке. // Укр. Биохим. Журн. 1984, Т. 54, с. 190-193.
31. Чухлова Э.А., Кудзина Л.Ю., Евтодиенко Ю.В. Влияние голодания на содержание и транспорт ионов калия в митохондриях печени крыс. // Укр. биохим. Журн., 1982, т. 54, № 2, с. 190-193.
32. Шортанова Т.Х., Шугалей В.С., Головина Т.Н. Особенности регуляции метаболизма у зимнеспящих. // Эволюционные аспекты гипобиоза и зимней спячки. 1986, с. 40-43.
33.Altshuld R., Hohl Ch., Ansel A., Brierley G. Compartmentation of K+ in isolated adult rat heart cells. // Arch. Biochem. Biophys., 1981, v. 209, № 1, pp 175-184.
34.Ammala C., Moorhouse A., Gribble F., Ashfield R., Proks P., Smith P.A., Sakura H., Coles B., Ashcroft F.M. Promiscous coupling between the sulfonylurea receptor and inwardly rectifying potassium channels. // Nature, 1996, 379:545-548.
35.Bagramyan K., Mnatsakanyan N., Trchounian A. Formate increases the F0F1-ATPase activity in Escherichia coli growing on glucose under anaerobic conditions at slightly alkaline pH. // Biochem. Biophys. Res. Commun., 2003, Jun 27, v. 306(2), p. 361-5.
36.Bernardi P., Azzone G.F. Electroneutral H+-K+ exchange in liver mitochondria. Regulation by membrane potential. // BBA, 1983, V. 724, pp 212-223.
37.Bogacka K., Nojtczak J. On the mechanism of mercurial-induced permeability of the mitochondrial membrane to K+. //FEBS Lett. 1979, V.100, pp 301-304.
38.Brierley G.P., Jurkowitz M., Jung D.W. Osmotic swelling of heart mitochondria in acetate and chloride salts. Evidence for two pathways for cation uptake. // Arch. Biochem. Biophys. 1978, V. 190, pp 181-192.
39.Brierley G.P. Monovalent cation transport by heart mitochondria. // Patology of cell membranes. 1983, №3, p 23-61.
40.Koster JC, Bentle KA, Nichols CG, Ho K. Assembly of ROMK1 (Kir 1.1a) inward rectifier K+ channel subunits involves multiple interaction sites. // Biophys J., 1998, Apr; 74(4), p 1821-1829.
41.Chavez E., Jung D.W., Brierley G.P. Energy-dependence exchange of K+ in heart mitochondria. K+ efflux. // Arch. Biochem. Biophys. 1977, V. 183, pp 460-470.
42.Clement J.P., Kunjilwar K., Gonzalez G., Schwanstecher M., Panten U., Aguilar-Bryan L., Bryan J. Assotiation and stoichiometry of K+-ATP channel subunita. // Neuron. 1997, v.18, p 827-838.
43.Dascal N, Schreibmayer W, Lim NF, Wang W, Chavkin C, DiMagno L, Labarca C, Kieffer BL, Gaveriaux-Ruff C, Trollinger D, et al. Atrial G protein-activated K+ channel: expression cloning and molecular properties. // Proc Natl Acad Sci USA., 1993, Nov 1; 90(21):10235-9.
44.Diwan J.J., Haley T., Sanadi D.R. Reconstitution of transmembrane K+- transport with a 53 kilodalton mitochondrial protein. // Biochem. Biophys. Res. ComMun., 1988, v. 31, p. 224-230.
45.Divan J. J., Tedeschi H. K+ fluxes mitochondrial membrane potential. // FEBS Lett. 1975, V.60, pp 176-179.
46.Garlid K.D. On the mechanism of regulation of the mitochondrial K+/H+ exchnger. // J. Biol. Chem. 1980, V. 255, pp 11273-11279.
47.Garlid K.D., Paucek P., Korcakova B., Woldegiorgis G. and Mironova G. Differential regulation of K flux through the reconstituted K-ATPchannels from cardiac mitochondria and sarcolemma. // ATP-sensitive K+ channels and sulfonylurea receptors. Houston, 1993, p. 81-85.
48.Garlid K.D., Paucek P., Yarov-Yarovoy V., Murrey H.N., Darbenzio R.B., D’Alonzo A.J., Lodge N.J., Smith M.A., Grover G.J. Cardioprotective effect of diazoxide and its interaction with mitochondrial ATP-sensitive K+ channels. Possible mechanism of cardioprotection.// Circ. Res., 1997 V 81, pp. 1072-1082.
49.Gomez-Puyou A., Tuena de Gomez-Puyou M. Monovalent cations in mitochondrial oxidative phosphorilation. // J. Bioenerg. Biomembr., 1977, v.9, № 1, pp 91-102.
50.Grigoriev, S.M., Skarga, Y.Y., Mironova, G.D. and Marinov, B.S. Regulation of mitochondrial KATP channel by redox agents. // Biochim Biophys Acta, 1999, v 1410, p 91-96.
51.Halestrap A.P., Davinson A.M. Inhibition of Ca2+-induced large-amplitude swelling of liver and heart mitochondria by cyclosporin is probably caused by the inhibitor binding to mitochondrial-matrix peptidyl-prolyl cis-trans isomerase and preventing it interacting with the adenine nucleotide translocase. // Biochem. J., 1990, V. 268, pp 153-160.
52.Halestrap A.P. The regulation of oxidation of fatty acids and other substrates in rat heart mitochondria by changes in the matrix volume induced by osmotic strength, valinomycin and Ca2+. // Biochem. J., 1987, V 244, pp159-164.
53.Ho K. The ROMK-cystic fibrosis transmembrane conductance regulator connection: new insights into the relationship between ROMK and cystic fibrosis transmembrane conductance regulator channels. // Curr Opin Nephrol Hypertens., 1998, Jan; v.7(1), p 49-58.
54.Inagaki N., Gonoi T., Clement J.P., Wang C.Z., Aguilar-Bryan L., Bryan J., Seino S. A family of sulfonylurea receptor determines the pharmacological properties of ATP-sensitive K+ channels. // Neuron. 1996, v.16, № 5, p 1011-1017.
55.Inoue I., Nagase H., Kishi K., and Higuti T. ATP-sensitive K+ channel in the mitochondrial inner membrane. // Nature, 1991, v 352, p. 244-247.
56.Yokoshiki H., Sunagava M., Seki T., Sperelakis N. ATP-sensitive K+-channels in pancreatic, cardiac, and vascular smooth muscle cells. //Am. J. Physiol., 1998, v. 274, p. 25-37.
57.Koster JC, Bentle KA, Nichols CG, Ho K. Assembly of ROMK1 (Kir 1.1a) inward rectifier K+ channel subunits involves multiple interaction sites. // Biophys J., 1998, Apr; 74(4), p 1821-1829.
58.Kraegeloh A, Amendt B, Kunte HJ. Potassium transport in a halophilic member of the bacteria domain: identification and characterization of the K+ uptake systems TrkH and TrkI from Halomonas elongata DSM 2581T. // J. Bacteriol. 2005, Feb; v. 187, № 3, p 1036-1043.
59.Kubo Y., Baldwin T.J, Jan YN, Jan L.Y. Primary structure and functional expression of a mouse inward rectifier potassium channel. // Nature, 1993, Mar 11, v. 362(6416):107-8.
60.Kubo Y., Reuveny E., Slesinger P.A., Jan Y.N., Jan L.Y. Primary structure and functional expression of a rat G-protein-coupled muscarinic potassium channel. // Nature, 1993, Aug 26, 364(6440): 758-9.
61.Liu Y., Sato T., O`Rourke B, Marban E. Circulation. Mitochondrial ATP-dependent potassium channels: novel effectors of cardioprotection? // Circulation, 1998, V. 97, pp. 2463-2469.
62.Mitchell P. Coupling of phosphorylation to electron and hydrogen transfer by a chemostatic type of mechanism. // Nature. 1961, V.191, pp 144-148.
63.Mitchell P., Moyle J. Respiration-driven proton translocation in rat liver mitochondria. // Biochem. J., 1971. V. 105, pp 1147-1162.
64.Mitchell P., Moyle J. Translocation of some anions, cations and acids in rat liver mitochondria. // Eur. J. Biochem., 1969, V.9, pp 149-155.
65.Paucek P., Mironova G., Mahdi F., Beavis A.D., Woldegiorgis G., and Garlid K.D. Reconstitution and partial purification of the glibenclamide-sensitive, ATP-dependent K+ channel from rat liver and beef heart mitochondria. // J. Biol. Chem, 1992, v 267, p. 26062-26069.
66.Rhoads D., Epstein W. Cation Transport in Escherichia coli. IX. Regulation of K+ transport. // J. of General Physiology, 1978, v. 72, p. 283-295.
67.Rottenberg H. The mechanism of energy-dependence ion transport in mitochondria. // J. Membr. Biol. 1973, V. 11, pp 117-137.
68.Scarpa A. Transport across mitochondrial membranes. // Membrane transport in biology. 1979, V. 2, pp 263-355.
69.Shears S.B., Brouk S.R. Ion transport difference in liver mitochondria from normal and thyroxine-treated rats. // J. Bioenerg. Biomembr., 1980, v. 12, № 5/6, p 379-393.
70.Shears S.B., Brouk S.R. The influence of thyroxine administered in vivo on the transmembrane protonic electrochemical potential difference in rat liver mitochondria. // Biochem. J., 1979, v. 178, № 3, p. 505-507.
71.Tedeschi H. The transport of cations in minochondria. // BBA. 1981, V.639, p 157-196.
72.Trchounian A, Kobayashi H. Kup is the major K+ uptake system in Escherichia coli upon hyper-osmotic stress at a low pH. // FEBS Lett, 1999, v. 447(23), p. 144-148.
73.Trchounian A, Kobayashi H. K+ uptake by fermenting Escherichia coli cells: pH dependent mode of the TrkA system operating. // Biosci. Rep., 2000, v. 20, № 4, p. 277-288.
74. Trchounian A.A., Ogandjanian E.S. An electrochemical study of energy-dependent potassium accumulation in E. coli. Part XIII. On the interaction of H+-ATPase complex F0F1 with Trk proteins in anaerobically grown cells. // Bioelectr. Bioenerg., 1992, v.27, p. 367-372.
0 комментариев