Молекулярные структуры, ответственные за транспорт калия в МХ
АТФ-ингибируемые калий-транспортирующие каналы
АТФ-чувствительный калиевый канал внутренней мембраны МХ
Модуляторы митоКАТФ канала
Механизмы защиты сердца при гипоксии, опосредованные активацией митоКАТФ
Феномен прерывистой гипобарической тренировки
Выделение МХ печени крысы
Подготовка белка с м.м. 55 кДа: выделение и очистка
Исследование ДНФ-индуцированного выхода К+ из митохондрий с помощью К+-селективного электрода
Очистка антител к АТФ-зависимому белку с м.м. 55 кДа на колонке с иммобилизованным Белком А
Изучение параметров АТФ-зависимого транспорта К+ в МХ печени сердца крыс с различной резистентностью к гипоксии
Изучение структурной организации митохондриального АТФ-зависимого калиевого канала
Навигация
АТФ-ингибируемые калий-транспортирующие каналы
Параметры функционирования митоКАТФ у животных с различной устойчивостью к гипоксии, а также у крыс, адаптированных к кислородному голоданию
118823
знака
4
таблицы
24
изображения
1.1.4 АТФ-ингибируемые калий-транспортирующие каналы
К+ каналы, активность которых ингибируется физиологическими концентрациями АТФ впервые были обнаружены в цитоплазматической мембране кардиомиоцитов [Noma, 1983]. С тех пор, аналогичные каналы были найдены в β клетках поджелудочной железы [Dunne and Petersen, 1991], в скелетной мускулатуре [Spruce et al., 1985; Woll et al., 1989], в нервных клетках [Jonas et al., 1990] и гладких мышцах [Standen et al., 1989]. Активность АТФ-чувствительных калиевых каналов (КАТФ каналов) связана с уровнем биоэнергетического метаболизма клетки (концентрацией АТФ) и электрическими свойствами возбудимости плазматической мембраны. Микромолярные концентрации АТФ и сульфонилмочевины – класс гипогликемических соединений, использующихся при лечении диабета, регулируют активность всех представителей семейства КАТФ каналов [Ashcroft et al., 1989]. Концентрация кальция и изменение мембранного потенциала практически не влияют на активность этих каналов [Ashcroft and Ashcroft, 1990]. В то же время, в кортикальных и гипоталамических нейронах КАТФ каналы [Ashford et al., 1989; Ashford et al., 1988] менее чувствительны к АТФ (для их ингибирования нужны миллимолярные концентрации). В эпителиальных клетках (назальные полипы) также были обнаружены КАТФ каналы с низкой чувствительностью к АТФ, но активирующиеся микромолярными концентрациями Ca2+ [Kunzelmann et al., 1989]. При этом, все вышеперечисленные каналы высокоселективны для ионов К+.
В МХ мембране также были обнаружены АТФ-зависимые К+ каналы. Так, в 1991 г. впервые с использованием метода петч-кламп [Inoue et al., 1991] на митопластах было показано, что во внутренней мембране МХ присутствуют высокоселективные по К+ каналы с невысокой проводимостью. Обнаружено, что каналы могут обратимо ингибироваться АТФ, а так же 4-аминопиридином и специфическим ингибитором цитоплазматического КАТФ канала – глибенкламидом с матриксной стороны [Inoue et al., 1991]. Однако, как было сказано выше, еще в 1981 году в лаборатории проф. Мироновой во внутренней мембране МХ был обнаружен К+-селективный АТФ-ингибируемый канал [Миронова и др., 1981; 1996; 1997; 1999], принадлежащий к семейству КАТФ каналов [Paucek et al., 1992]. Данный канал имеет те же характеристики проводимости, что и канал, обнаруженный методом пэтч-кламп. В настоящее время он интенсивно изучается, поскольку играет важную роль в функционировании клетки. Однако вопрос о его структуре требует дальнейших исследований.
1.2 АТФ-зависимый калиевый канал цитоплазматической мембраны
К настоящему времени КАТФ каналах плазматической мембраны хорошо изучены. Изначально их классификация велась по величине проводимости одиночных каналов [Ashcroft and Ashcroft, 1990], а позднее, с развитием молекулярно-биологических методик, по аминокислотному составу белковой молекулы канала [Yokoshiki et al., 1998].
1.2.1 Структурная организация цитоплазматического АТФ-зависимого калиевого канала
Молекулярно-биологические и электрофизиологические исследования последних лет позволили определить структуру цитоКАТФ канала. Исследуемый канал состоит из двух белков: KIR – inward rectifying K+ channels, формирующий пору канала, и SUR – sulphonyl urea receptor, регуляторная субъединица, придающая каналу чувствительность к модуляторам. SUR-субъединица содержит нуклеотидсвязывающие участки, локализующиеся на цитоплазматической стороне, а также участки связывающие активаторы калиевых каналов (potassium channel activators, PCOs) и MgАДФ. Каналы подсемейства KIR6.0 сами обладают чувствительностью к АТФ. Роль SUR заключается, вероятно, в облегчении доступа АТФ к АТФ-связывающему участку KIR [Tucker et al., 1997; Yokoshiki et al., 1998]. В настоящее время механизмы молекулярного взаимодействия KIR и SUR окончательно не выяснены.
Результаты исследования электрофизиологических свойств нативных КАТФ каналов позволяют предположить, что они принадлежат к классу калиевых каналов внутреннего выпрямления (KIR). В 1993 году на многие KIR каналы (KIR1.1 [Ho et al., 1993], KIR2.1 [Kubo et al., 1993], KIR3.1 [Dascal et al.,1993]) были получены клоны. Их выделили в отдельное подсемейство KIR-каналов (KIR1.0-6.0), основанное на унифицированной номенклатуре [Chandy and Gutman, 1993]. Гидрофобные участки клонированных KIR каналов содержат два мембранопересекающих сегмента М1 и М2, образующих пору. Подобно потенциалзависимым К+ каналам, KIR каналы, вероятно, формируют тетрамер [Doupnik et al., 1995; Salkoff and Jegla, 1995; Nichols et al., 1996; Nichols and Lopatin, 1997].
Также был клоирован регуляторный белок, содержащий участок, связывающий сульфонилочевину (SU-связывающий сайт) [Aguillar-Bryan et al., 1995; Thomas et al., 1995; Inagaki et al., 1996; Isomoto et al., 1996]. Клонированные регуляторные белки получили название SUR. SUR содержит два нуклеотидсвязывающих участка (NFBs) с цитоплазматической стороны. N-конец находится с внешней стороны мембраны и содержит два домена, предположительно из 9 и 4 пронизывающих мембрану спиралей (Рис.2) [Yokoshiki et al., 1998].
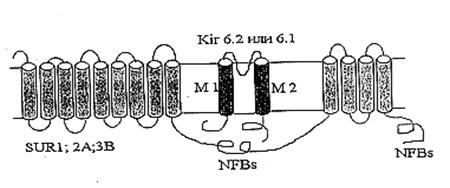
Рис. 2. Схематическое изображение цитоплазматического КАТФ канала как комплекса KIR и SUR [Yokoshiki H. et al., 1998]
Считается, что нативный КАТФ канал представляет собой комплекс из четырех KIR6.x и четырех SUR субъединиц [Clement et al., 1997]. Различные типы регуляторных субъединиц, SUR1, SUR2A и SUR2B, обнаружены, соответственно, в поджелудочной железе, сердце и гладких мышцах [Ammala et al., 1996; Inagaki et al., 1996; Yamada et al., 1997].
0 комментариев