Навигация
Фосфолипазы, их классификация и свойства
58905
знаков
0
таблиц
7
изображений
Содержание
Введение
1. Фосфолипазы
1.1 Классификация. Свойства
1.2 Система фосфолипаза С - инозитол-3-фосфат
2. Фосфолипазы А2
2.1 Общие сведения (реакция, открытие, строение)
2.2 Классификация и свойства
2.2.1 Цитозольные ФЛА2
2.2.2 Секреторные ФЛА2
2.2.3 Кальцийнезависимая ФЛА2
2.3 Субстратная специфичность
2.4 Ингибиторы ФЛА2
2.4.1 Неконкурентное ингибирование
2.4.2 Конкурентное ингибирование
2.5 Значение для организма при нарушении активности
2.6 Использование ФЛА2 в медицине
2.7 Биологическая роль ФЛА2
Список литературы
Введение
Фосфолипазы (англ. phospholipase) ферменты класса гидролаз, катализирующие гидролиз фосфоглицеридов.. В зависимости от положения гидролизуемой связи в фосфолипиде различают 4 основных класса фосфолипаз: A, B, C и D.
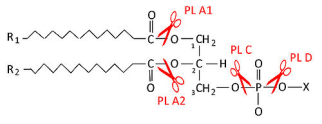
Лизофосфолипиды расщепляются под действием фосфолипаз L (существование позиционно специфичных фосфолипаз L1 и L2 не доказано). Фосфолипазы В - устаревшее назв. препаратов, обладающих активностью по типу фосфолипаз А и L.
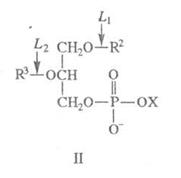
X - остаток холина, серина, миоинозита и др.; для фосфолипаз L1 R2=C(O)R4, R3=H; для фосфолипаз L2 R2=H, R3=C(O)R4
Каждое из семейств фосфолипаз неоднородно и включает ферменты, значительно отличающиеся по молекулярным массам, субъединичному составу и другим свойствам. Все фосфолипазы наиболее активно катализируют гидролиз на поверхности раздела фаз фосфолипид - вода; медленно гидролизуют водорастворимые субстраты.
Фосфолипаза A1 - (КФ 3.1.1.32, англ. phospholipase A1) отщепляет SN-1 ацильную цепь.
Фосфолипаза A2 - (КФ 3.1.1.4, англ. phospholipase A2) отщепляет SN-2 ацильную цепь.
Фосфолипаза B -(лизофосфолипаза, англ. phospholipase B) отщепляет обе SN-1 и SN-2 ацильные цепи. Фосфолипаза, обладающая активностями как фосфолипазы А1 так и А2, то есть способной гидролизовать ацильную цепь фосфолипида в sn-1 и sn-2 положениях.
Фосфолипаза C - (КФ 3.1.4.3, англ. phospholipase C) гидролизует связь между глицериновым остатком фосфолипида и полярной фосфатной группой, при этом образуются диацилглицерин и фосфат-содержащая полярная группа.
Фосфолипаза D - (КФ 3.1.4.4, англ. phospholipase D) гидролизует связь между фоасфатной группой и спиртовой группой, при этом высвобождаются фосфатидная кислота и спирт. Существует 2 изоформы этой фосфолипазы D1 и D2.
Фосфолипазы играют важную роль в обмене липидов в живых организмах. Их используют для определения структуры фосфоглицеридов и места их локализации в мембранах.
1. Фосфолипазы.
1.1 Классификация. Свойства
Фактически различают несколько фосфолипаз группы А, они являются составной частью многих тканей и секретов живых организмов.
Фосфолипазы A1 в большинстве своем - внутриклеточные ферменты, часто мембраносвязанные, не нуждаются в коферменте. Их молекулярные массы варьируют в пределах 15-90 тыс.; оптимальная каталитическая активность проявляется при рН 4,0 (для лизосомальных ферментов) или 8,0-9,5 (для ферментов микросом, плазматических мембран и цитозоля); широко распространены в животных тканях (печень, сердце, мозг) и в микроорганизмах (Bacillus subtilis, В. megateiium, Mycobacter phlei, Escherichia coli).
Фосфолипазы А1 отщепляют ацильную цепь фосфолипида в sn-1 положении. При действии фосфолипазы А1 на фосфолипид образуется 1-лизофосфолипид и жирная кислота. Фосфолипаза является активным компонентом змеиного яда гемолитического действия.
Фосфолипазы A2 - наиболее изученные представители фосфолипаз. Известны 3 группы фосфолипаз A2: 1) ферменты ядов змей, рептилий и насекомых, существующие в виде большого количества изоформ; 2) ферменты поджелудочной железы млекопитающих, продуцирующиеся в организме в виде зимогенов (предшественников с большей молекулярной массой) и активирующиеся трипсином; 3) внутриклеточные ферменты из крови и тканей животных, среди которых имеются как растворимые, так и мембраносвязанные.
Фосфолипазы A2 первых двух подгрупп являются водорастворимыми ферментами с молеклярной масссой 11-19 тыс. (некоторые активны в виде димеров), обладают высокой стабильностью благодаря большому числу (6-7) дисульфидных связей. Оптимальная каталитическая активность при рН 7,5-9,0; рI от 4,0 до 10,5; кофермент - Ca2+. Для множества представителей этих подгрупп фосфолипаз известны первичная и пространственная структур. В активном центре обнаружены остатки гистидина и аспарагиновой кислоты. Свойства внутриклеточных фосфолипаз A2 (третья подгруппа) зависят от субклеточной локализации фермента. Их молекулярная масса 12-75 тыс.; оптимальная каталитическая активность при рН 4,2-9,0. Некоторые ферменты этой подгруппы не содержат коферментов.
Фосфолипазы В выделены из растений, микроорганизмов, яда пчел, тканей млекопитающих. Ферменты этой группы крайне неспецифичны, катализируют гидролиз различных сложноэфирных связей, обладают литическим (разрушающим) действием по отношению к биологически мембранам (что обусловливает их токсичность). Молекулярная масса фосфолипаз В 15-65 тыс., они менее стабильны, чем фосфолипазы А; их оптимальная каталитическая активность проявляется при рН от 4,5 (лизосомальный фермент) до 10,0 (ферменты ядов). Фосфолипазы не имеют коферментов, не ингибируются этилендиаминтетрауксусной кислотой. Некоторые фосфолипазы В ингибируются диизопропилфторфосфатом и п-хлормеркурбензойной кислотой. Универсальные ингибиторы для всех фосфолипаз В - ПАВ.
Фосфолипаза В способна гидролизовать ацильную цепь фосфолипида в sn-1 и sn-2 положениях Как правило фосфолипаза действует на лизолецитин (лизофосфатидилхолин), образующийся в результате действия фосфолипазы А1 на лецитин (фосфатидилхолин).
Фосфолипазы С обнаружены у бактерий Clostridium, Bacillus и Pseudomonas, а также в клетках млекопитающих (печень, мозг, поджелудочная железа). Для некоторых из них характерна строгая специфичность по отношению к спиртовой группе молекулы субстрата, например к остатку холина (фосфолипазы Cx) и миоинозита (фосфолипазы Си). Молекулярная масса фосфолипаз С от 23 до 51 тыс. Ионы Zn2+ являются для них коферментом и стабилизатором. Оптимальная каталитическая активность при рН около 7 для фосфолипаз Cx, и при рН < 7 для фосфолипаз Си.
Фосфолипаза С, гидролизующая фосфодиэфирную связь между глицериновым остатком фосфолипида и полярной фосфатной группой, относится к фосфодиэстеразам также как и фосфолипаза D. Фосфолипаза С является ключевым ферментом метаболизма фосфатидилинозитола и липидных сигнальных путей.
Фосфолипаза С активируется Gαq или Gβγ субъединицами G-белка. Таким образом, она является частью G-белоксвязанного рецептора (англ. G protein-coupled receptor) и соответствующего сигнального пути или частью трансмембранного рецептора с внутренней или ассоциированной тирозинкиназной активностью.
Фосфолипаза С гидролизует фосфатидилинозитол (PIP2) на два вторичных медиатора инозитолтрифосфат (IP3) и диацилглицерин (DAG). Эти медиаторы становятся вовлечены в последующие этапы сигнальных путей. В частности, они модулируют кальциевые каналы эндоплазматического ретикулума и протеинкиназу С, соответственно.
Фосфолипаза D относится к группе важных ферментов, которые в живых системах выполняют разнообразные функции от усвоения питательных веществ до синтеза биологически активных соединений. Обнаружены в растениях (овощи, водоросли), микроорганизмах и в тканях животных. Их молекулярная масса 90-116 тыс. Оптимальная каталитическая активность при рН 4,7-8,0. Катионные ПАВ ингибируют фосфолипазы D, анионные - активируют.
Фосфолипаза D проявляет прежде всего гидролитическую активность, в результате которой происходит расщепление сложноэфирной связи между остатком фосфатидной кислоты и спирта в молекулах фосфолипидов (ФЛ). При этом последний замещается на водород, но возможен перенос остатка фосфатидной кислоты на самые разные гидроксилсодержащие акцепторы, что представляет большой интерес для биотехнологии, так как трансфосфатидилирующая активность фосфолипазы может быть использована для синтеза разнообразных лекарственных препаратов.
Фосфолипаза D специфически расщепляет фосфатидилхолин на фосфатидную кислоту и холин, высвобождая последний в цитоплазму.
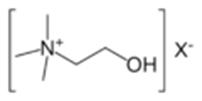
фосфатидилхолин фосфатидная кислота холин
Похожие работы
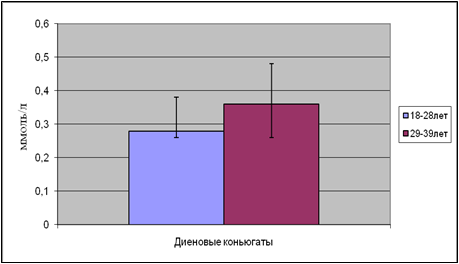
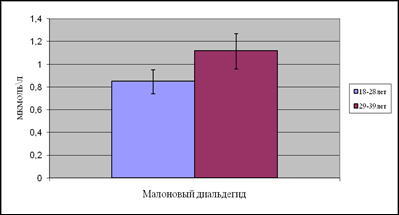
... находится на невысоком уровне, но при стрессовых ситуациях происходит усиленная активация процессов ПОЛ под действием АФК, что приводит к патологическому состоянию. 3.1. Содержание продуктов перекисного окисления липидов в плазме крови небеременных женщин. В данной работе было определено содержание диеновых коньюгатов и малонового диальдегида у небеременных женщин (N=31), которые составили ...
... фосфолипиды. На основе проведенного анализа литературы можно предположить, что диапазон функциональных возможностей мезотелия обусловлен состоянием микроциркуляции. Становится очевидным, что для определения потери антиадгезивных свойств мезотелием брюшины, необходимо изучить морфологические пределы нормального функционирования клеток мезотелия в условиях динамизма микроциркуляциии. Главный ...
... , оранжевыми миндалинами и аккумуляцией эфиров ХС в других ретикулоэндотелиальных тканях. Патология связана с ускоренным катаболизмом апо А-I [8, 2000]. Переваривание и всасывание липидов. Желчь. Значение. На заре формирования современного учения о внешнесекреторной функции печени, когда естествоиспытатели располагали лишь первыми ...
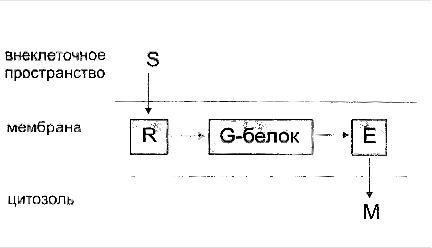
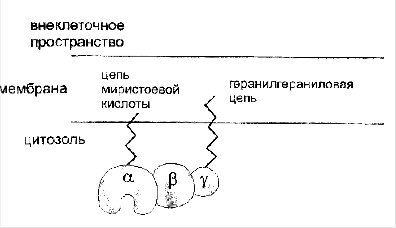
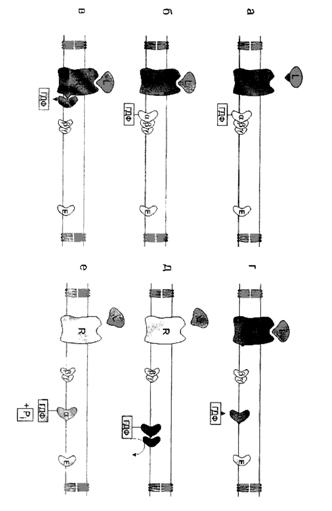
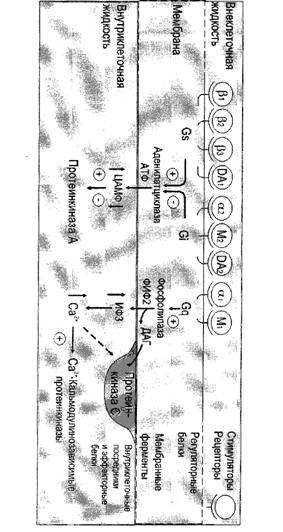
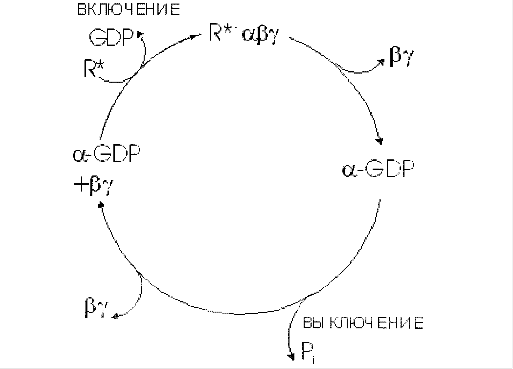
... сигнала по инозитолфосфатному пути. Протеинкиназы Все полярные сигнальные молекулы, действующие на клетку-мишень через мембранные рецепторы, осуществляют свою биологическую функцию путём фосфорилирования специфических белков и ферментов, регулирующих метаболизм в клетке. Фосфорилирование изменяет (увеличивает или уменьшает) их активность. Катализируют фосфорилирование белков (протеинов) ...
0 комментариев