Навигация
2.2.1 Секреторные ФЛА2
Секреторные ФЛА2 имеют молекулярную массу около 14 кДа, характеризуются абсолютной необходимостью ионов Са2+ в миллимолярных концентрациях для каталитической активности, рН оптимум находится в интервале 7-9.
В настоящее время описано десять типов секреторной ФЛА2 (IВ, IIА, IIC, IID, IIE,IIF,III,V, X,XII) которые различаются первичной структурой и расположением дисульфидных мостиков. Все типы секреторных фосфолипаз представляют собой глобулярные белки, богатые цистеином (5-8 дисульфидных мостиков), что обеспечивает стабильность фермента, в том числе устойчивость к протеолизу и денатурации. Фермент не проявляет избирательности в отношении жирнокислотного состава фосфолипидов, но предпочтительно гидролизует отрицательно заряженные фосфолипиды (фосфатидную кислоту и фосфатидилглицерол).
Долгое время была известна только одна ФЛА2, которая в изобилии присутствует в панкреатической жидкости (тип IB). В 1989 г. была открыта и клонирована фосфолипаза типа IIA, которая хранится в секреторных гранулах тромбоцитов и концентрация которой значительно увеличивается в местах воспаления, таких как синовиальная жидкость при ревматоидном артрите. Эти белки имеют большое сходство с белками яда змей. Среди белков ядов пчёл и ящериц были обнаружены другие фосфолипазы, которые отнесены к типу III. У млекопитающих белок, соответствующий этому типу, был обнаружен только в 2000 г.. Новый период в исследованиях секреторных фосфолипаз начался в 1994 г., когда были открыты белки типа IIC и V. Это открытие привело к пересмотру роли этого семейства белков в регуляции функций клеток и интенсивному поиску новых аналогичных белков. Были открыты белки типа X, IID, IIE и IIF, XII.
Все эти белки (кроме белка типа III) имеют молекулярную массу 14-17 кДа, содержат гистидин в каталитическом центре и проявляют фосфолипазную ктивность в присутствии миллимолярных концентраций кальция. Они имеют высококонсервативные аминокислотные последовательности в каталитическом участке (DXCCXXHD) и участке кальцийсвязывающей петли (XCGXGG), а также 6-8 консервативных сульфидных мостиков. В активном центре находится также аспартат, который совместно с кальцийсвязывающей петлей выполняет роль кармана для иона Ca2+.
Ферменты этого семейства не проявляют специфичности ни по отношению к группе, связанной с остатком фосфатидной кислоты, ни по отношению к ацильной группе в положении sn-2. Наличие в фосфолипидах окисленных форм жирных кислот увеличивает активность секреторных фосфолипаз, что предполагает их участие в регуляции вязкости мембран при окислительном стрессе. Из данных рентгеноструктурного анализа первых двух групп белков следует существование гидрофобного канала, в который входит молекула фосфолипида после межфазного связывания белка на фосфолипидной поверхности.
Секреторная ФЛА2 конститутивно содержится в различных клетках, участвующих в развитии иммунных и воспалительных ответов: макрофагах, тучных клетках, фибробластах и тканях таких органов, как печень, селезенка, тимус, костный мозг, кишечник. Активность фермента на уровне клеток регулируется за счет его индукции различными воспалительными стимулами (интерлейкин-1 и интерлейкин-6, фактор некроза опухоли,липополисахарид, интерферон-g, форболовые эфиры, фактор роста нервов). В соответствии с индукцией разнообразными стимуляторами промотер гена IIA содержит нуклеотидные последовательности TATA и CAAT, а также последовательности связывания таких факторов транскрипции, как AP-1, C/EBP, CREB, NF-kB, STAT, PPA Rg. В некоторых клетках экспрессия ФЛА2 зависит от предварительной активации цитозольной фосфолипазы PLА2a, при этом предполагается вовлечение в процесс регуляции фосфолипазы IIA продуктов 12/15- липоксигеназного пути. Глюкокортикоиды (стероидные противовоспалительные препараты) являются супрессорами экспрессии фосфолипазы IIA.
Изменение экспрессии фосфолипазы IIA во многих случаях связано с модуляцией простагландиновой ветви каскада арахидоновой кислоты. Так, при стимуляции интерлейкином-1 и фактором некроза опухоли активировался как синтез и секреция фосфолипазы, так и синтез простагландина Е2 и простациклина в мезангиальных или эндотелиальных клетках, соответственно. При добавлении к клеткам антител к фосфолипазе синтез простациклина частично подавлялся. Из результатов, полученных в последние годы, следует, что секреторные фосфолипазы (IIA и схожие с ней V, X) участвуют в процессах как быстрой, так и замедленной продукции арахидоновой кислоты и простаноидов.
Следует отметить, что добавление секреторных фосфолипаз во внеклеточную среду приводит к активному высвобождению арахидоновой кислоты и синтезу простаноидов активированными клетками, но практически не влияет на гидролиз фосфолипидов мембран покоящихся клеток.
Помимо роли фермента, ответственного за наличие арахидоновой кислоты, секреторные фосфолипазы могут выступать в роли физиологически активных веществ. Так, в тучных клетках специфические ингибиторы секреторных фосфолипаз уменьшали стимулированную фактором роста нервов экспрессию циклооксигеназы-2. При этом каталитически неактивные мутанты белка фосфолипазы также были способны вызывать экспрессию циклооксигеназы-2, т.е. этот эффект не зависит от ферментативных свойств фосфолипазы. Механизм этого процесса не ясен. Возможно, вовлекаются функции секреторной ФЛА2 как лиганда специфических рецепторов.
Действительно, в 1995 г. было показано, что существуют специфические белки, которые связываются с фосфолипазой типа IB (константа диссоциации образовавшегося комплекса 1 нмоль/л) и проявляют различные биологические эффекты. За последующие 10 лет обнаружено ещё много белков, растворимых и мембраносвязанных, которые способны связывать секреторную фосфолипазу. Однако свойства «классического» рецептора, который при связывании лиганда активирует систему внутриклеточной передачи сигнала, проявляет только один из этих белков. Это белок M-типа, или sPLA2R. Ген данного белка локализован во второй хромосоме; белковая последовательность имеет 75 % гомологии среди ряда видов млекопитающих; ген имеет 1 копию и ничем не похож на другие гены. Белок имеет молекулярную массу 180-200 кДа, значительная его часть расположена во внеклеточной области, в цитозоле находится последовательность из 40 аминокислотных остатков. Белок без этого цитозольного участка встречается в растворимой форме и показана его роль как ингибитора эффектов секреторных фосфолипаз. У человека белок экспрессирован в поджелудочной железе, лёгких и почках. Показана важная роль активации этого рецептора в развитии эндотоксического шока.
На рисунке представлена схема участия рецептора секреторной фосфолипазы в реализации биологической роли фермента на уровне клеток.

На схеме показано, как активные формы фосфолипазы sPLA2-IB или sPLA2–X проявляют свою ферментативную активность, что приводит к появлению липидных медиаторов, а также являются высокоаффинными лигандами для рецептора, локализованного в плазматической мембране. Взаимодействие фосфолипазы с рецептором мембраны приводит к индукции митогенактивированных протеинкиназ (MAPK) и соответствующего пути внутриклеточного проведения сигнала, что стимулирует различные ответы клеток: пролиферацию и миграцию клеток, синтез ими физиологически активных веществ. При потере контакта с мембраной рецептор сохраняет возможность связывать фосфолипазу, что позволяет регулировать активность последней как фермента и как лиганда.
Таким образом, секреторные фосфолипазы играют существенную роль в развитии и распространении воспалительных процессов в организме. Экспрессия секреторных фосфолипаз значительно увеличивается при разнообразных воспалительных заболеваниях. По этой причине разрабатывают селективные ингибиторы ферментативной активности белков этого семейства как потенциально новый класс противовоспалительных веществ. Для поиска ингибиторов секреторных фосфолипаз перспективно применение методов компьютерного моделирования, так как эти низкомолекулярные белки (их молекулярная масса около 14 кДа) получены в кристаллическом состоянии, известны их трёхмерные структуры. Есть основания полагать, что в ближайшие 5-10 лет на основании результатов этих исследований будут созданы новые терапевтические технологии и новые лекарственные средства.
Похожие работы
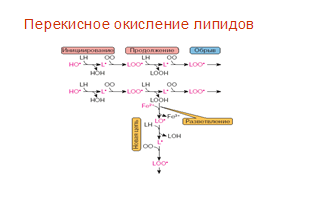
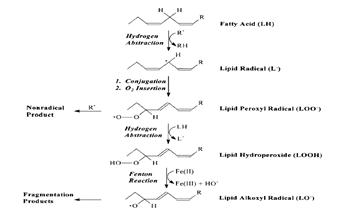
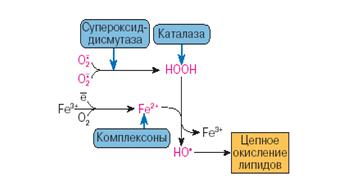
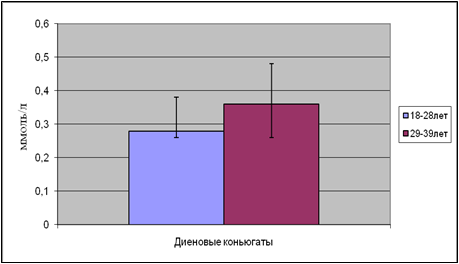
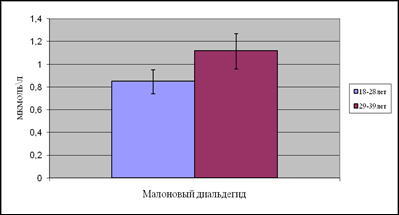
... находится на невысоком уровне, но при стрессовых ситуациях происходит усиленная активация процессов ПОЛ под действием АФК, что приводит к патологическому состоянию. 3.1. Содержание продуктов перекисного окисления липидов в плазме крови небеременных женщин. В данной работе было определено содержание диеновых коньюгатов и малонового диальдегида у небеременных женщин (N=31), которые составили ...
... фосфолипиды. На основе проведенного анализа литературы можно предположить, что диапазон функциональных возможностей мезотелия обусловлен состоянием микроциркуляции. Становится очевидным, что для определения потери антиадгезивных свойств мезотелием брюшины, необходимо изучить морфологические пределы нормального функционирования клеток мезотелия в условиях динамизма микроциркуляциии. Главный ...
... , оранжевыми миндалинами и аккумуляцией эфиров ХС в других ретикулоэндотелиальных тканях. Патология связана с ускоренным катаболизмом апо А-I [8, 2000]. Переваривание и всасывание липидов. Желчь. Значение. На заре формирования современного учения о внешнесекреторной функции печени, когда естествоиспытатели располагали лишь первыми ...
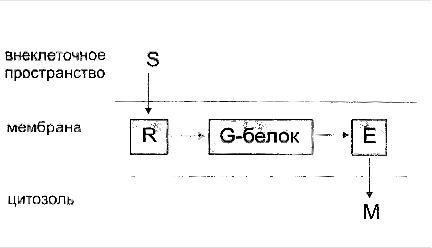
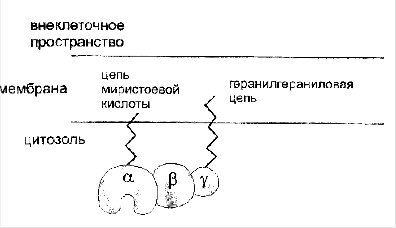
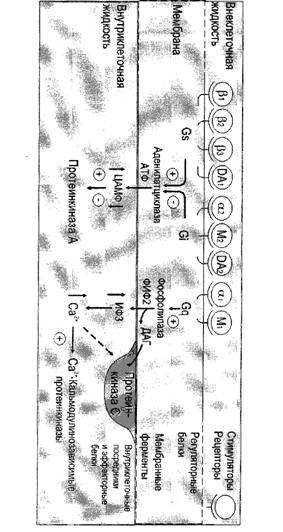
... сигнала по инозитолфосфатному пути. Протеинкиназы Все полярные сигнальные молекулы, действующие на клетку-мишень через мембранные рецепторы, осуществляют свою биологическую функцию путём фосфорилирования специфических белков и ферментов, регулирующих метаболизм в клетке. Фосфорилирование изменяет (увеличивает или уменьшает) их активность. Катализируют фосфорилирование белков (протеинов) ...
0 комментариев