Введение
Менее жестко запрограммированная эволюция
Выводы
Осложнения
Хромосомные и химические основы наследственности
Связь классической и молекулярной генетики
Неодарвинизм и популяционная генетика
Холдейн, Фишер и значение отбора
Сьюэлл Райт, адаптивные ландшафты и случай
Рекомендуемая литература
Меланизм у бабочек — подход a posteriori
Приложение теории игр к поведению животных
Изменчивость в природных популяциях; ее значение для адаптации
Адаптации на благо группы и адаптации групп
Адаптации самого полового размножения как такового и порожденные им адаптации
Рекомендуемая литература
Канализация развития
Заключения
Кладизм (5, 7)
Эволюционная систематика (13)
Как образуются таксоны — неодарвинистская точка зрения, или синтетическая теория эволюции
Альтернативная точка зрения — прерывистая эволюция
Сравнение теорий постепенной и прерывистой эволюции
Заключения
Навигация
Рекомендуемая литература
Принципы эволюции
242666
знаков
12
таблиц
37
изображений
2.5. Рекомендуемая литература.
Можно назвать ряд хороших сводок по современной генетике: Гейл (7), Айала и Киджер (1), Берри (2). В этих работах используются почти исключительно коэффициенты отбора, тогда как изложение Риклефса (9) основывается На мальтузианском параметре (см. также Кук (4).) Несколько сложнее учебники Раухгардена (10) и Чарлзуорта (3), в которых рассматриваются популяции с перекрывающимися поколениями. Наиболее ясным изложением количественной генетики и, в частности, наследуемости (/i2) все еще остается книга Фальконера (6). С молекулярной генетикой можно познакомиться в книгах Уотсона (12) и Вудса (15). История генетики кратко и интересно изложена Лудовичи (8), а крайне субъективное, но тем не менее интересное описание бурного развития молекулярной генетики принадлежит Уотсону (13). Молекулярная генетика развивается так стремительно, что очень трудно уследить за всеми происходящими в ней событиями. Оригинальное изложение ее развития, дающее определенное представление об увлекательности предмета и о ряде важных его аспектов, можно найти в работе Уотсона и Туза 114).
Глава 3. АДАПТАЦИЯ.
3.1. Введение.
В гл. 2 мы определили приспособленность, исходя из относительного распространения генов в генофондах, то есть исходя из эффекта естественного отбора. Однако связь между распространением данного гена и самим геном зависит от того, как он взаимодействует со своей средой, а это взаимодействие он осуществляет не непосредственно, а через находящегося под генным контролем посредника — через фенотип. Поэтому полезно рассмотреть, почему в данных экологических условиях некоторые фенотипы способствуют передаче генов лучше, чем другие (более приспособлены), иными словами, рассмотреть причину отбора. Подобный подход носит название адаптационистской программы и сочетает в себе рассмотрение формы и функции фенотипов наряду с их экологией и эволюцией.
Со времен Дарвина такая адаптационистская программа всегда пользовалась популярностью у биологов, однако центр тяжести ее постоянно смещался. Как указывал, например, Доукинс (12, 13), дарвинисты пытались рассматривать признаки с точки зрения их полезности для выживания и размножения отдельных особей (то, что Доукинс называет подходом с позиций эгоистичного организма), тогда как после неодарвинистского переворота главное внимание стали уделять объяснению признаков с точки зрения их полезности для переноса генетических единиц (то есть генов и сцепленных групп генов). Этот новый взгляд на вещи привел к тому, что Доукинс называет подходом с позиций эгоистичного гена. При этом фенотипические признаки должно оценивать лишь с учетом того, какое влияние они могут оказывать на распространение тех генов, которые их контролируют. Наконец, признаки организма иногда оценивают в свете того, какую пользу они могут принести той группе (например, семье, популяции, виду), в которую входит данный организм. Такой подход можно назвать подходом с позиций неэгоистичного организма.
Большинство активных приверженцев адаптационизма придерживаются подхода с позиций эгоистичного организма, поскольку, по их мнению, такой подход более или менее совместим с концепцией эгоистичного гена, то есть рассматривает фенотипические признаки с точки зрения их значения для выживания и размножения организмов — носителей этих признаков, что в свою очередь должно быть связано положительной корреляцией с распространением генов, кодирующих эти фенотипические признаки. Дальнейшие объяснения этой методологии и соответствующие примеры будут приведены в разд. 3.2—3.5. Подход с позиций неэгоистичного организма рассматривается в разд. 3.6. Проблемы, в изучении которых были использованы оба подхода— с позиций эгоистичного и неэгоистичного организма,— обсуждаются в разд. 3.7; и, наконец, в разд. 3.8 мы вернемся к разбору взаимоотношений между эгоистичным геном и эгоистичным организмом.
3.2. Методология.
Существуют два различных, но, как станет очевидным, взаимозависимых пути, по которым развивалась адаптационистская программа (5).
Метод 1 (апостериорный!сравнительный подход). При этом методе изучают и сравнивают признаки у близких видов в разные периоды истории данной группы (то есть по ископаемым остаткам) или же — в современной фауне — у видов, занимающих различные экологические ниши. Затем (после такого изучения) дается объяснение обнаруженных различий в адаптивном плане. В отношении различий, коррелированных со временем, принимается допущение, что изменения в признаках следуют за изменениями экологических условий и (или) представляют собой прогрессивные адаптивные усовершенствования. Так, например, направления в эволюции лошадей (рис. 3.1) объясняют переходом их от жизни в лесах к жизни на равнинах. В процессе эволюции наблюдается: 1) увеличение общих размеров, в частности длины конечностей, которые при этом становятся и более стройными; 2) уменьшение числа пальцев до одного и разрастание «ногтя» этого оставшегося пальца с образованием копыта; 3) увеличение резцов в ширину и коренных зубов в высоту вместе с развитием сложных складок твердого цемента на их жевательной поверхности. Изменения 1 и 2 можно рассматривать как адаптации к быстрому передвижению по твердой почве, а изменение 3 — как адаптацию к питанию жесткой степной травой.
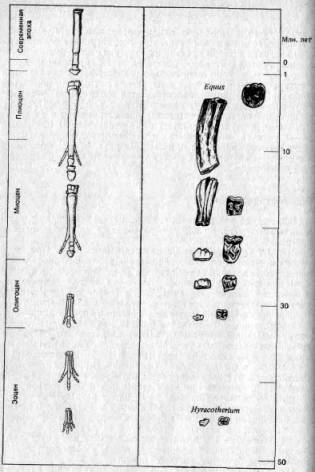
Рис. 3.1. Направления эволюции конечностей и коренных зубов у лошадей (объяснения см. в тексте).
Что касается изменений, коррелированных с пространством, то считается, что дивергенция признаков у популяций, принадлежащих к одному и тому же или к близким видам, произошла в результате различных давлений отбора, испытываемых этими популяциями в тех различных экологических условиях, в которых они находятся. Следовательно, нам важно иметь возможность точно определять различия в тех экологических условиях, под действием которых находятся организмы! Физиологическим экологам, изучающим прибрежную фауну, известно множество примеров близкородственных видов, обитающих в разных частях приливно-отливной зоны и поэтому в разной степени подверженных обсыханию в промежутке между двумя приливами и воздействию волн. Морфологические, физиологические и поведенческие различия между видами, обитающими в разных частях приливно-отливной зоны, во многих случаях можно объяснить как адаптации к этим экологическим различиям; один из таких примеров приведен в табл. 3.1. В этом случае большую устойчивость к высоким температурам и иссушающим условиям у Patella vulgata, обитающей в верхней части этой зоны, чем у P. aspera, обитающей в ее нижней части, можно рассматривать как приспособление к более частому и более длительному пребыванию в воздушной среде в промежутке между приливами. Кроме того, более низкую скорость потери воды в воздушной среде у P. vulgata по сравнению с P. aspera можно отнести за счет различий в форме раковины у этих двух видов (у P. vulgata поверхность, обсыхающая в воздушной среде, меньше); таким образом, морфологические различия поддаются объяснению с позиций адаптационизма.
Таблица 3.1. Некоторые аспекты физиологической экологии морских видов блюдечка (по данным Девиса (10,11))
| Признак | patella vulgata | Patella aspera |
| Зона обитания Выживаемость при t<30°C Скорость потери воды тканями Способность противостоять потере воды | Верхняя Высокая Низкая Высокая | Нижняя Низкая Высокая Низкая |
| Форма раковины | | |
В связи с методом 1 возникает ряд проблем.
1. Не во всех случаях можно твердо установить, являются ли наблюдаемые различия генотипическими или лишь фенотипиче-скими. Так, например, различия в форме раковины, приведенные в табл. 3.1, можно отнести за счет прямого воздействия волн, то есть в этом случае, если бы оба вида развивались в одних и тех же условиях, никаких различий между ними не было бы. Подобного рода негенетические различия (называемые иногда фенотипической пластичностью) в данном контексте интереса не представляют. Отметим, однако, что фенотипическая пластичность может быть детерминирована генетически, а может быть адаптивной. Примеры исследований, в которых был проведен более точный генетический анализ, будут даны в разд. 3.3.
2. Наличие корреляции само по себе нельзя считать доказательством существования причинно-следственной зависимости. Возможно, что в возникновении различий участвуют какие-то скрытые переменные, играющие более важную роль. Возможна также, что различия в форме раковины у блюдечка обусловлены различиями в выеданий хищниками, а это последнее регулируется воздействием волн или обсыханием в промежутке между приливами. В таком случае различия в признаках следовало бы объяснять в свете того, в какой степени раковины определенной формы обеспечивают защиту от хищников, а не от обсыхания или воздействия волн.
3. Не все различия обязательно должны быть адаптивными. Некоторые из них могли возникнуть случайно (дрейф, эффект основателя). Другие могли возникнуть не в результате прямого .отбора, а как «побочные эффекты» признаков, обладающих селективным преимуществом, например в случаях плейотропии (один ген имеет несколько фенотипических эффектов, причем не все они должны быть адаптивными) или аллометрии (см. разд. 4.2). Одним из таких признаков может быть старение. Его нельзя назвать явно адаптивным, поскольку оно, по определению, снижает жизнеспособность и повышает уязвимость затронутых им организмов. Старение может быть адаптивным применительно к группе в целом, например, в том смысле, что оно освобождает место для новых мутаций, связанных с молодыми особями, которые замещают старых особей и, следовательно, стимулируют эволюцию и долговременное выживание группы, однако такую точку зрения с позиций группового отбора разделить трудно (см. разд. 3.6).
В отличие от этого несколько других теорий связывают старение с плейотропными эффектами. Суть их сводится к следующему: в природе организмы гибнут по большей части не от старости, а от несчастных случаев, болезней и хищников. В результате вмешательства этих внешних факторов вероятность выживания часто с возрастом снижается, так что существует некий возраст, после которого животные обычно гибнут и не вносят никакого репродуктивного вклада в популяцию. Если один ген обладает несколькими эффектами, среди которых есть как благоприятные, так и неблагоприятные, то отбор будет направлен на то, чтобы задержать, проявление последних до того периода жизненного цикла, до которого организмы обычно не доживают. Проявление этих гибельных эффектов (внутренне присущие организму факторы смертности) становится заметным лишь тогда, когда внешние факторы смертности устранены, как в лаборатории или в человеческом обществе.- С этой точки зрения старение — не какая-то особая адаптация, а «побочный эффект» генов, которые обладают независимыми от старения благоприятными эффектами, проявляющимися на более ранних стадиях жизненного цикла.
4. Признаки могут быть адаптивными и могут быть обусловлены отбором, но тем не менее различия между ними не обязательно должны иметь селективную основу. Иными словами, для решения одних и тех же проблем могут развиваться различные адаптации, то есть в адаптивном ландшафте имеются многочисленные адаптивные пики, и какой из них займет популяция, зависит от ее начального состава. Это может лежать в основе различия между индийским носорогом, имеющим один рог, и африканским носорогом, у которого два рога. Рога — приспособление для защиты от хищников, и вряд ли можно думать, что один рог — специфическое приспособление для индийских условий, а два — для африканских. Более вероятно, что изначально индийская и африканская популяции несколько различались, в частности по своим морфогенетическим системам, а поэтому под действием одних и тех же селективных сил они достигли различных адаптивных пиков (25).
Метод 2 (априорный/предсказательный подход). Этот метод используют не для объяснения корреляций между фенотипичес-кими признаками и экологической изменчивостью, а для того, чтобы предсказать, какими должны быть эти корреляции. В принципе он позволяет делать предсказания до проведения наблюдений. Подобного рода априорные эволюционные рассуждения часто бывают основаны на допущении, что естественный отбор — процесс оптимизирующий, то есть что в некотором смысле он ведет к эволюции наилучших из возможных признаков. Инженеры и экономисты, которые также стремятся избрать наилучшие решения для конкретных технических или экономических проблем, разработали специальные методы, например теорию оптимального управления, и эти методы находят также применение и в биологии (34). При использовании принципа оптимальности для решения любой проблемы должны быть выполнены следующие основные требования: 1) все возможные решения данной задачи должны быть известны; 2) каждому решению должно быть возможно приписать некоторые числа или сложные математические функции, соответствующие либо стоимости (и), либо цене (с) этого решения относительно какого-либо заранее заданного условия. Математическая задача принципа оптимальности состоит в том, чтобы среди значений v и с найти максимальное v или минимальное с.
Основное условие применимости принципа оптимальности состоит в том, чтобы прибыль и цену можно было измерить, а это зависит в свою очередь от ясного недвусмысленного определения того, какие требования предъявляются к системе. У инженеров и экономистов всегда есть заранее установленные критерии, с которыми можно сопоставлять поведение изучаемой системы. Согласно теории неодарвинизма (см. разд. 3.1), фенотипические особенности биологических систем должны быть такими, чтобы максимизировать расселение потомков (несущих данный признак), генов, а быть может, даже генных комплексов, так что прибыль и цену можно оценивать относительно этих требований. Если бы мы могли точно определять признаки в зависимости от их воздействия на выживание, время генерации и репродуктивный вклад, то было бы относительно легко выбрать те из них, которые максимизируют неодарвинистскую приспособленность. К сожалению, непосредственно вычислить эту величину обычно бывает возможно только для нескольких признаков, и приходится довольствоваться лишь косвенными допущениями. Поэтому весь метод сводится к следующему: 1) допускаем, что отбор максимизирует неодарвинистскую приспособленность (основная гипотеза); 2) переводим 1 в фенотипическую меру приспособленности (вспомогательная гипотеза); 3) используя соответствующие математические методы, находим признак, который максимизирует 2 (или минимизирует ее снижение); 4) сравниваем это предсказание с тем, что наблюдается в природе или обнаруживается в специально созданных экспериментальных условиях. В этой программе адаптационисты редко пытаются опровергнуть основную гипотезу. Они обычно исходят из допущения, что эта гипотеза более или менее верна, а затем пытаются уточнить свое понимание эволюции фенотипа, критически оценивая вспомогательные гипотезы.
Кон (7, 8), а позднее Милсум и Роберж (31) изучали адаптацию применительно к кровеносной системе позвоночных. Они хотели выяснить, какой ширине сосудов будет благоприятствовать естественный отбор. Конечно, оценить диаметр кровеносных сосудов исходя из его воздействия на выживание, скорость развития и размножения не представляется возможным, а поэтому необходима была какая-то вспомогательная гипотеза; такой гипотезой послужило допущение, что отбор должен благоприятствовать самой экономичной структуре. Оно было основано на том, что, чем меньше ресурсов будет затрачено на построение и поддержание кровеносной системы, тем больше их останется у организма для борьбы с болезнями и хищниками и для того, чтобы производить потомство. Кон и его соавторы рассматривали две оценочные функции (рис. 3.2): одна из них возрастает с уменьшением диаметра сосудов (потому что сопротивление трения потоку жидкости при этом возрастает и делает необходимым большую затрату энергии сердцем на прокачивание крови), а другая возрастает с увеличением диаметра (потому что на построение и поддержание системы больших размеров необходимо больше ресурсов). Вспомогательная гипотеза требует, чтобы отбор благоприятствовал той системе, которая обходится дешевле; именно это решение может быть получено графически (рис. 3.2) или из уравнений, определяющих оценочные функции.
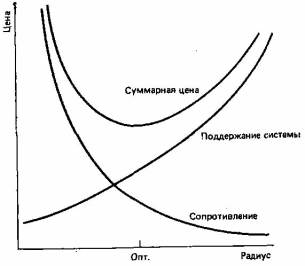
Рис. 3.2. Модель Кона, предсказывающая оптимальный радиус (Опт.) для кровеносных сосудов. Существуют две оценочные функции, одна из которых возрастает, а другая уменьшается с увеличением радиуса сосуда (см. текст). Оптимальным является тот радиус, который минимизирует сумму значений этих функций, то есть представляет собой наиболее экономичное решение.
Подставив в эти уравнения реальные числа, можно вычислить конкретные оптимальные диаметры и сравнить их с результатами наблюдений (табл. 3.2). Кон и его соавторы получили достаточно хорошее совпадение между реальными и ожидаемыми диаметрами. Следует, однако, помнить, что своей работой они не пытались опровергнуть неодарвинизм; они лишь хотели понять, что он означает применительно к развитию кровеносной системы. Данная вспомогательная гипотеза позволяет сформулировать некое онтогенетическое правило, которое может оказаться применимым и для других систем, а именно: системы органов должны быть возможно более экономичными по форме и функции.
Таблица 3.2. Предсказания Кона и результаты измерений реальных сосудов.
| Предсказания | Результаты измерений | |
| Радиус аорты, см | 0,43 | 0,5 |
| Диаметр капилляров, мкм | 2,2 | 4,0 |
Метод 2 также сопряжен с некоторыми затруднениями. Главные из них относятся к формулировке ряда допущенных решений, из которых можно было бы выбрать оптимальное. С одной стороны, мы не можем сделать этого, не прибегая к реальным наблюдениям, то есть метод 2 строится на основе метода 1, а это означает, что модели создаются в соответствии с фактическими данными, а не предсказывают их. С другой стороны, не все мыслимые решения реально осуществимы, а поэтому необходимо некоторым образом ограничить их. Иными словами, оптимизационные методы можно рассматривать как поиски на адаптивном ландшафте (см. разд. 2.3.3) самых высоких пиков, в условиях, когда доступны не все пики: некоторые пики могут соответствовать таким фенотипам, возникновение которых невозможно по морфогенетическим причинам, другие могут оказаться исключенными вследствие исходного генетического состава популяции до начала действия отбора. Возможно также, что уровень генетической изменчивости в популяции окажется недостаточным для достижения наивысшего пика. К сожалению, установить a priori эти ограничения чрезвычайно трудно, и применение метода 2 может дать о них столько же информации, сколько и о самой адаптации.
Похожие работы
... матери требовали максимальной "притирки" плода к условиям организма матери, поэтому генетический контроль должен был быть минимальным. 2. Рост мозга. Развитие коры головного мозга. Развитие интеллекта. Эмоциональное развитие. 2.1 Функциональные блоки мозга Дифференциация систем мозговой коры происходит постепенно, и это приводит к неравномерному созреванию отдельных мозговых структур, ...
... . Кроме того, ДНК имеется в митохондриях, которые передаются только по материнской линии. Исследования мутаций в этих ДНК позволяют реконструировать историю биологического развития человечества, происхождение отдельных рас и народностей. Мутация может быть рецессивной, доминантной и полудоминантной в зависимости от состояния гена, в котором она произошла. Гены мутируют с определенной частотой, и ...
— концепция «2К» (кооперация и коммуникация) — эта концепция была разработана в 1900 г. русским ученым-философом П.А.Кропоткиным (1842—1921). В ней идет речь в основном о механизме биологической эволюции. . Термин «эволюция» (лат. evolutio — развертывание) в науке о живом представляет фундаментальное понятие для объяснения возникновения и развития всего живого. Эволюция подразумевает необратимое ...
... о противоречивом пути развития синтетической теории эволюции, о преодолении кризисов в научном познании, направленном на более полное и глубокое понимание закономерностей развития органического мира. Важно отметить, что основы научной теории эволюции заложил Ч. Дарвин. Как господствующее эволюционное учение дарвинизм существовал с 1859 до 1900 гг., т.е. до переоткрытия законов Г. Менделя. До ...
0 комментариев