Введение
Менее жестко запрограммированная эволюция
Выводы
Осложнения
Хромосомные и химические основы наследственности
Связь классической и молекулярной генетики
Неодарвинизм и популяционная генетика
Холдейн, Фишер и значение отбора
Сьюэлл Райт, адаптивные ландшафты и случай
Рекомендуемая литература
Меланизм у бабочек — подход a posteriori
Приложение теории игр к поведению животных
Изменчивость в природных популяциях; ее значение для адаптации
Адаптации на благо группы и адаптации групп
Адаптации самого полового размножения как такового и порожденные им адаптации
Рекомендуемая литература
Канализация развития
Заключения
Кладизм (5, 7)
Эволюционная систематика (13)
Как образуются таксоны — неодарвинистская точка зрения, или синтетическая теория эволюции
Альтернативная точка зрения — прерывистая эволюция
Сравнение теорий постепенной и прерывистой эволюции
Заключения
Навигация
Хромосомные и химические основы наследственности
Принципы эволюции
242666
знаков
12
таблиц
37
изображений
2.2. Хромосомные и химические основы наследственности.
2.2.1. Цитологические основы наследственности.
За последние три десятилетия XIX в. в изучении клеточной основы жизни были достигнуты выдающиеся успехи. Ряд исследователей, главным образом немецких, открыли, что гаметы представляют собой клетки и что клетка — это не просто комочек однородного вещества, а некая структура, состоящая из ядра и цитоплазмы. Благодаря усовершенствованию микроскопов и разработке специальных методов окрашивания стало возможным более детально рассмотреть ядро. Оказалось, что оно также неоднородно: в нем были обнаружены нитевидные структуры, состоящие из вещества, названного хроматином; соответственно структуры эти получили название хромосом. Мнение о том, что хромосомы — подходящие кандидаты на роль материальных носителей наследственности, впервые высказал Август Вейсман (August Weismann, 1834—1914). Он представлял себе наследственность как передачу материала — того, что он называл зародышевой плазмой, — от родителей потомкам и предполагал, что зародышевая плазма, сохраняя непрерывность и обособленность, проходит через весь онтогенез данного индивидуума и «выплескивается» в следующее поколение, покидая сому — свое прежнее тело. Эти взгляды Вейсмана были основаны на его наблюдениях, которые показывали, что у многих животных образование половых клеток и их обособление от соматических тканей происходит на очень ранней стадии эмбрионального развития. Отсюда проводившееся им разграничение между зародышевой плазмой и сомой. В своей «Эволюционной теории», вышедшей в 1903 г., Вейсман еще более сузил границы зародышевой плазмы и свел ее к ядру половых клеток, а затем — к хромосомам и хроматину (впервые эти термины были предложены В. Вальдейером в 1888 г.). Он это сделал потому, что, судя по гистологическим данным, хромосомы вели себя именно так, как им следовало себя вести, если бы они представляли собой вещество наследственности: они удваивались и разделялись на две равные группы при делении соматических клеток; число их уменьшалось вдвое при образовании гамет, предшествующем смешиванию мужского и женского вкладов во время оплодотворения. В настоящее время мы называем эти два процесса соответственно митозом и мейозом.
Итак, на основе всех полученных данных менделевские факторы получили название генов (термин, введенный В. Л. Иоган-сеном в 1888 г.), которые, как предположили исследователи, находятся в хромосомах. Эта теория, конечно, коренным образом отличалась от теории слитной наследственности, которой придерживался Дарвин. Зародышевая линия была отделена от сомы, а тем самым и от приобретенных признаков. Гены дискретны и не сливаются. Кроме того, хромосомная теория позволяла лучше осмыслить данные некоторых экспериментов по скрещиванию, например данные Моргана по сцеплению у дрозофилы (см. разд. 2.1.2).
2.2.2. Химические основы наследственности.
Локализация генов в хромосомах была установлена, однако не все биологи были так уверены, как Вейсман, в их чисто химической природе. Сам Т. Г. Морган в своей книге «Механизм менделевской наследственности» (Т. Н. Morgan, A. H. Sturte-vant, H. J. Muller, С. В. Bridges, Mechanism of Mendelian Heredity, H. Holt and Co., 1915) писал: «Предположение, что частицы хроматина, неотличимые одна от другой и почти гомогенные при исследовании любыми известными методами, могут в силу своей материальной природы наделять всеми свойствами жизни, превосходит воображение даже самого убежденного материалиста».
Создание разнообразия, или, иными словами, генетической информации, из единообразия —вот в чем заключался главный вопрос, а ответ на него состоял в том, что хроматин оказался не таким гомогенным, каким он представлялся. Данные хими-деского анализа показали, что хроматин — это нуклеиновая кислота, состоящая из повторяющихся субъединиц (нуклеоти-дов), соединенных друг с другом связями, в образовании которых участвуют остатки либо рибозы (рибонуклеиновые кислоты, РНК), либо дезоксирибозы (дезоксирибонуклеиновые кислоты, ДНК).
Крупнейший шаг вперед в этой области был сделан в середине пятидесятых годов нашего века, когда Дж. Д. Уотсон (James D. Watson) и Фрэнсис Крик (Francis Crick) расшифровали строение ДНК на основе химических данных в сочетании с рентгеновскими «снимками» ее молекулы. Каждая нуклеиновая кислота состоит из двух цепей, соединенных химическими связями, так что структура в целом имеет вид веревочной лестницы. Перекладинами этой лестницы служат связи между нуклеотидами, причем нуклеотиды расположены таким образом, что против определенной большой единицы может находиться только одна определенная малая единица. Поэтому говорят, что две стороны лестницы комплементарны. Эта лестница свернута наподобие гибкого электрического провода, образуя ныне знаменитую двойную спираль (см. рис. 2.6).
Вслед за созданием модели структуры ДНК был высказан ряд соображений относительно ее функций. Во-первых, последовательность оснований в молекуле ДНК может быть различной и в этой последовательности может быть закодирована генетическая информация. Хроматин оказался негомогенным. Во-вторых, комплементарность означает, что если отделить две стороны «веревочной лестницы» одну от другой, то каждая из них могла бы служить матрицей для построения другой. Следовательно, информация может точно и безошибочно реплицироваться и передаваться от одной клетки другой и от родителей потомкам (рис. 2,6,Г).
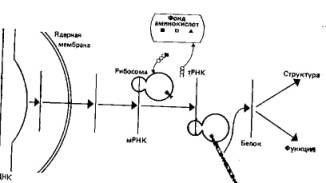
Рис. 2.7. Транскрибирование ДНК в мРНК и трансляция в белки. Трансляция происходит на субклеточных органеллах — рибосомах; молекулы транспортной РНК (тРНК) выбирают из клеточного фонда аминокислот нужную аминокислоту и переносят ее на соответствующее место на матрице. Образующийся таким образом белок может затем выполнять в клетке структурные или ферментативные функции. (Calow P., Biological Machines, Edward Arnold Publ., London, 1976.)
Наконец, спиральная структура означает, что эта жизненно важная генетическая информация защищена от превратностей внешнего мира. Время от времени в процессе репликации возникают случайные нарушения, ведущие к тому, что в классической генетике называли мутациями. Подобные события должны быть редкими и могут вызываться внешними факторами (например, облучением), но они не направлены на удовлетворение каких-либо определенных «потребностей».
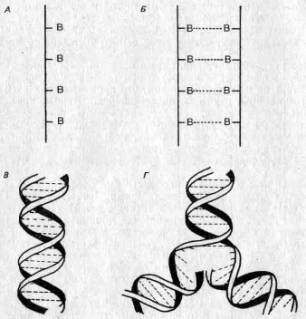
Рис. 2.6. Строение ДНК. А. Нуклеотиды (В), соединенные в цепь. Б. Комплементарные цепи, образующие «веревочную лестницу» при помощи водородных связей (прерывистые линии). В, «Веревочная лестница», свернутая в двойную спираль. Г. Репликация: цепи раскручиваются, и каждая из них служит матрицей для образования комплементарной цепи.
Все эти положения, разработанные Уотсоном и Криком, оказались в основном верными и вызвали бурное развитие исследований по молекулярной генетике. Было известно, что ядро и нуклеиновые кислоты играют ключевую роль в синтезе белка, а поскольку белки принимают важнейшее и непосредственное участие в построении клетки (в виде строительных блоков) и в регуляции ее функций (в виде ферментов), то нетрудно видеть, каким образом нуклеиновые кислоты могли бы определять признаки организма. Нуклеиновые кислоты состоят из повторяющихся субъединиц — нуклеотидов, а белки состоят из аминокислот; поэтому можно было допустить, что последовательность нуклеотидов кодирует последовательность аминокислот. Поскольку, однако, аминокислот различных типов в белках больше, чем нуклеотидов в нуклеиновых кислотах (соотношение их составляет примерно 5:1), одна аминокислота не может кодироваться только одним нуклеотидом. Теоретически было нетрудно показать, что лишь группы из трех нуклеотидов (триплеты) могут обеспечить достаточное разнообразие (43, или 64 альтернативные комбинации), а вскоре существование три-плетного кода было установлено экспериментально. У эукариот перенос генетической информации при синтезе белка происходит в два этапа: от дезоксирибонуклеиновых кислот (ДНК) к рибонуклеиновым кислотам (РНК)—процесс, известный под названием транскрипции, и от РНК к белкам — трансляция. Упрощенная схема переноса генетической информации представлена на рис. 2.7.
Похожие работы
... матери требовали максимальной "притирки" плода к условиям организма матери, поэтому генетический контроль должен был быть минимальным. 2. Рост мозга. Развитие коры головного мозга. Развитие интеллекта. Эмоциональное развитие. 2.1 Функциональные блоки мозга Дифференциация систем мозговой коры происходит постепенно, и это приводит к неравномерному созреванию отдельных мозговых структур, ...
... . Кроме того, ДНК имеется в митохондриях, которые передаются только по материнской линии. Исследования мутаций в этих ДНК позволяют реконструировать историю биологического развития человечества, происхождение отдельных рас и народностей. Мутация может быть рецессивной, доминантной и полудоминантной в зависимости от состояния гена, в котором она произошла. Гены мутируют с определенной частотой, и ...
— концепция «2К» (кооперация и коммуникация) — эта концепция была разработана в 1900 г. русским ученым-философом П.А.Кропоткиным (1842—1921). В ней идет речь в основном о механизме биологической эволюции. . Термин «эволюция» (лат. evolutio — развертывание) в науке о живом представляет фундаментальное понятие для объяснения возникновения и развития всего живого. Эволюция подразумевает необратимое ...
... о противоречивом пути развития синтетической теории эволюции, о преодолении кризисов в научном познании, направленном на более полное и глубокое понимание закономерностей развития органического мира. Важно отметить, что основы научной теории эволюции заложил Ч. Дарвин. Как господствующее эволюционное учение дарвинизм существовал с 1859 до 1900 гг., т.е. до переоткрытия законов Г. Менделя. До ...
0 комментариев