Введение
Менее жестко запрограммированная эволюция
Выводы
Осложнения
Хромосомные и химические основы наследственности
Связь классической и молекулярной генетики
Неодарвинизм и популяционная генетика
Холдейн, Фишер и значение отбора
Сьюэлл Райт, адаптивные ландшафты и случай
Рекомендуемая литература
Меланизм у бабочек — подход a posteriori
Приложение теории игр к поведению животных
Изменчивость в природных популяциях; ее значение для адаптации
Адаптации на благо группы и адаптации групп
Адаптации самого полового размножения как такового и порожденные им адаптации
Рекомендуемая литература
Канализация развития
Заключения
Кладизм (5, 7)
Эволюционная систематика (13)
Как образуются таксоны — неодарвинистская точка зрения, или синтетическая теория эволюции
Альтернативная точка зрения — прерывистая эволюция
Сравнение теорий постепенной и прерывистой эволюции
Заключения
Навигация
Осложнения
Принципы эволюции
242666
знаков
12
таблиц
37
изображений
2.1.2. Осложнения.
Как и можно было ожидать, дальнейшие исследования показали, что генетический механизм сложнее, чем представлял себе Мендель. Тем не менее ни одно из этих осложнений не опровергает основных допущений Менделя о дискретности генов и их большой стабильности. Ниже кратко изложены некоторые наиболее значительные осложнения.
1. Доминирование. Оперение кур андалузской породы отливает синим, но среди цыплят всегда попадаются черные особи и особи с белыми пятнами. На самом деле андалузки гетерозиготны по окраске оперения, но черная окраска не полностью доминирует над пятнистой. Это не может служить доводом в пользу слитной наследственности, потому что в потомстве от скрещиваний между гетерозиготными особями наблюдается выщепле-ние пятнистой и черной окраски в соответствии с менделевски-ми соотношениями.
Теперь мы знаем, что степень доминирования одного гена над другим зависит от его экспрессивности, а не от каких-либо существенных различий между генетическими системами. При кодоминантности оба аллеля экспрессируются в одинаковой мере (то есть аддитивны), именно это и происходит у кур андалузской породы. При частичном, или неполном, доминировании один из аллелей не активен (не экспрессируется), но другой активен полностью, так что в фенотипе эффект проявляется наполовину. При полном доминировании (то, что наблюдал Мендель) эффект одного аллеля эквивалентен эффекту двух аллелей, а при сверхдоминировании эффект одного аллеля выражен в большей степени, чем эффект двух таких аллелей в гомозигот-ном состоянии. Картина еще больше осложняется в тех случаях, когда экспрессия аллелей, находящихся в одной части (локусе) генного набора, регулируется экспрессией аллелей, находящихся в других локусах. Это явление известно под названием эпис-таза. Например, при скрещивании двух гомозиготных мышей, одна из которых доминантна по окраске шерсти — агути (серая шкурка), а другая — рецессивна (черная шкурка), все потомство должно было бы иметь окраску агути. Большая часть мышат действительно относится к типу агути, однако встречаются и альбиносы, причем частота их выше той, которую можно былом бы отнести за счет мутаций. Происходит это потому, что на ок-" раску шкурки влияет еще и другая пара генов, находящихся в другом локусе. Доминантный аллель в этом локусе определяет нормальную окраску, детерминированную аллелями главного локуса, а рецессивный аллель определяет альбинизм, и любая мышь, гомозиготная по этому аллелю, будет альбиносом независимо от состояния главного локуса.
2. Сцепление. В экспериментах по гибридизации, проводившихся Менделем, факторы, которые он изучал, распределялись независимо, однако теперь мы знаем, что гены расположены в хромосомах (см. разд. 2.2.1) и поэтому распределение каждого из них по гаметам зависит от их относительного распределения по хромосомам. Если два локуса находятся в одной и той же хромосоме и притом расположены близко один от другого, то они будут передаваться вместе.
Например, Томас Хант Морган (Thomas Hunt Morgan), положивший в 1909 г. начало генетическому изучению плодовой мушки (Drosophila), обнаружил у двух генов D. melanogaster отклонение от второго закона Менделя; один из этих генов определяет цвет глаз (рг — пурпуровый, рг+— красный), а дру-. гой — длину крыла (vg —зачаточное, vg+ — нормальное). Обратите внимание на то, что принцип обозначения генов здесь несколько иной: обозначение получает мутантный (обычно рецессивный) аллель, а доминантный аллель отличают от него индексом +. Такая система обозначений была принята потому, что для каждого локуса дрозофилы обнаружено большое число мутантных аллелей; она до сих пор используется генетиками, работающими с дрозофилой, наряду с другими обозначениями (см. (!)). Морган скрещивал гетерозиготных самок (ргрг+ vgvg+) с гомозиготными самцами (prpr vgvg). На основании классической менделевской генетики следовало ожидать, что в fi все четыре возможные сочетания генов будут представлены в равных количествах: prpr vgvg, prpr vgvg+, prpr+ vgvg, prpn v§v§+ (рис. 2.3, А). Однако на самом деле частота родительских форм (prpr+ vgvg+ и prpr vgvg) в потомстве была в 10 раз выше частоты двух других форм. Морган объяснил это, допустив, что два родительских аллеля сцеплены один с другим и поэтому передаются в одной родительской хромосоме, как это показано на рис. 2.3, Б. Но если эти два гена сцеплены, то как вообще могли возникнуть генные комбинации prpr vgvg+ и prpr+ vgvg? Очевидно, генетический материал каким-то образом переносится с одной хромосомы на другую (рис. 2.3, В), и существование такого явления, названного кроссинговером, было впоследствии подтверждено цитологическими наблюдениями. Ясно, что чем ближе друг к другу расположены два локуса в хромосоме, тем выше вероятность их совместной передачи и тем менее вероятен их кроссинговер. Таким образом, по частоте сцепления между локусами можно судить об их относительном местоположении в хромосомах, что служит основой для составления карт хромосом.

Рис. 2.3. Интерпретация механизма наследования цвета глаз и формы крыла у дрозофилы (объяснения см. в тексте).
Первым признаком, хромосомную локализацию которого Удалось установить (в результате ряда ключевых экспериментов, проведенных главным образом в США между 1900 и 1910 гг.), был пол. У млекопитающих и у дрозофилы одна хромосома (назовем ее хромосомой X) имеется у особей женского пола в двух экземплярах, а у особей мужского пола — только в одном. Вместо второй Х-хромосомы у самцов имеется отличная от нее Y-хромосома. Таким образом, самки несут хромосомы XX и образующиеся у них яйцеклетки содержат одну Х-хросому, а самцы несут хромосомы XY и образующиеся у них сперматозоиды содержат либо Х, либо Y-хромосому. Если допустить, что находящиеся в этих хромосомах ген или гены контролируют пол, то становится возможным объяснить наблюдаемое соотношение полов 1 : 1 при помощи следующей простой схемы:
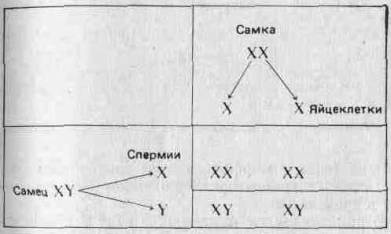
(Отметим, что у птиц и у некоторых насекомых наблюдается обратная картина: гетерогаметны самки, а самцы гомогаметны.) Соотношение полов рассматривается более подробно в разд. 3.7. Половые хромосомы X и Y морфологически отличаются от остальных хромосом (аутосом) и друг от друга. Поэтому можно ожидать, что они содержат уникальные аллели, которые не могут переходить из одной хромосомы в другую в результате кроссинговера. Это может привести к отклонениям от предсказаний Менделя; явление это известно под названием сцепления с полом. Например, аллель, определяющий красный цвет глаз У Дрозофилы, о котором говорилось выше, как и аллель белого Цвета глаз, локализован в Х-хромосомах. В Y-хромосомах этих аллелей нет. Красный цвет глаз доминирует над белым. Когда Морган скрещивал красноглазых самок с белоглазыми самцами, все потомство fj оказалось красноглазым. При скрещивании красноглазых самцов и самок из fi он получил красноглазых и белоглазых мух в соотношении 3:1, но все белоглазые мухи были мужского пола. Отношение красноглазых самок к красноглазым самцам составляло 2:1. Если, заменив обозначения, принятые на стр. 27, обозначить аллель красных глаз через Xw, а аллель белых глаз — через Xw, то нетрудно дать объяснение! наблюдаемым результатам:
Рациональное объяснение этого дает рис. 2.5. Вообще чем больше генов участвует в определении данного признака, тем более непрерывна изменчивость по этому признаку.
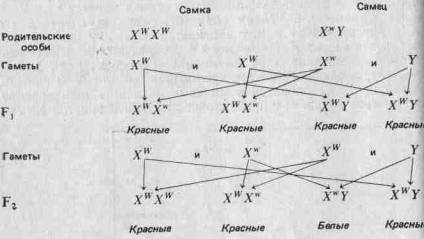
Такая ассоциация генов с морфологически различными хромосомами служит очень хорошим подтверждением того, что гены организованы в хромосомы.
3. Непрерывно изменяющиеся признаки. Мендель сконцентрировал внимание на признаках, резко отличающихся один от другого, однако большинству признаков свойственна непрерывная изменчивость. Так, например, организмы по общим размерам обычно не распадаются на дискретные классы, а образуют непрерывный ряд от минимальных до максимальных для данной группы размеров. Кроме того, чаще всего особи распределяются по данному признаку таким образом, что большая их часть концентрируется вблизи среднего значения признака, а по обе стороны от него частота постепенно сходит на нет (см., например, рис. 2.4). В течение некоторого времени полагали, что эта изменчивость выходит за рамки менделевской генетики; У. Ф. Уэлдон (W. F. Weldon) примерно в 1900 г. даже подверг критике данные самого Менделя, поскольку при более тщательном анализе оказалось, что большая часть так называемых дискретных признаков горошка подвержена значительной изменчивости (например, степень морщинистости семян и интенсивность их зеленой окраски). Однако в 1909 г. X. Нильсен-Эле (Н. Nilssen-Ehle) показал, что менделевские факторы могут контролировать количественные признаки, если один такой признак детерминируется большим числом генов (полигены), каждый из которых обладает небольшим, аддитивным действием (см. стр. 26).
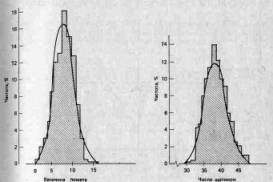
Рис. 2.4. Частотное распределение величины помета у мышей и числа щетинок на вентральной поверхности брюшка у дрозофилы. В обоих случаях кривая распределения приближается к нормальной (гауссовой) кривой. (Futuyama D. J., Evolutionary Biology, Sinauer Associates, Massachusetts, 1979.)
Помимо генетической основы изменчивости признаков существует еще и влияние окружающих условий. Например, на общие размеры животного или растения решающее влияние оказывают те условия, в которых оно развивается. Возможно, все признаки организма представляют собой продукт воздействия генетических и средовых факторов, так что нормальные распределения, подобные изображенным на рис. 2.4, — результат сочетания этих факторов. Передается по наследству только та компонента этой изменчивости, которая обусловлена генами; ее выражают как Долю общей изменчивости и называют наследуемостью (часто обозначается как h2). Измерение наследуемости имеет важное значение для селекционеров и животноводов; ее можно определить по соотношению между признаками родительских особей и потомков, выращенных в тех же условиях.
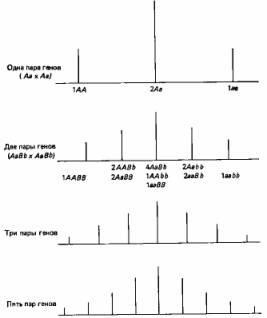
Рис. 2.5. Схема, иллюстрирующая возникновение фенотипической изменчивости при корпускулярной наследственности.
Допустим, что «сила» данного признака зависит от соотношения доминантных и рецессивных аллелей. В таком случае, чем больше число расщепляющихся локусов, определяющих этот признак, тем больше число возможных сочетаний доминантных и рецессивных признаков, а тем самым и число возможных фенотипических классов. (Strickberger M. W., Genetics, Macmillan, New York, 1968.)
Вильгельм Людвиг Иогансен (W. L. Johansen), специалист по генетике растений, был одним из первых ученых, обратив- J ших внимание (примерно в 1900 г.) на то, что каждый организм— продукт среды, в которой он обитает, и генов, которые он содержит. На этой основе Иогансен различал фенотип, то есть, организм, который мы видим, и генотип — совокупность содержащихся в нем генов, и это проведенное им разграничение сохранило свое важное значение по сей день.
4. Мейотшеский драйв. Аллели, кодирующие один и тот же признак, находятся в отдельных хромосомах (гомологичных хромосомах), так что каждая клетка данной родительской особи содержит обычно некоторое число пар гомологичных хромосом (диплоидное состояние). При формировании гамет гомологичные-хромосомы должны быть отделены одна от другой, с тем чтобы каждая гамета в конечном итоге получила по одной хромосоме от каждой пары (гаплоидное состояние). Этот процесс разделения известен под названием мейоза и лежит в основе первого закона Менделя — закона расщепления. Мейоз обеспечивает формирование гетерозиготной особью одинакового числа гамет, содержащих разные аллели. Однако некоторые процессы, детерминируемые генами, способны нарушить эти результаты. Такие процессы могут быть обусловлены событиями, как предшествующими мейозу, так и следующими за ним; и те и другие приводят к так называемому мейотическому драйву. Обзор по этой проблеме см. (16).
Один интересный пример связан с так называемыми генами SD (segregation distorter), обнаруженными у дрозофилы (5). В этом случае гомологичные хромосомы конъюгируют, но гены SD оказывают на своих партнеров такое воздействие, что те в дальнейшем образуют нежизнеспособные сперматозоиды. Существование подобных SD-генов проявляется более четко в тех случаях, когда они редки, потому что при этом у них больше шансов оказаться в гетерозиготном состоянии. По мере того как они становятся более обычными, конъюгация между ними происходит чаще и они разрушаются. Поэтому следует ожидать, что гены SD сбалансированы в популяциях на довольно низких уровнях.
Похожие работы
... матери требовали максимальной "притирки" плода к условиям организма матери, поэтому генетический контроль должен был быть минимальным. 2. Рост мозга. Развитие коры головного мозга. Развитие интеллекта. Эмоциональное развитие. 2.1 Функциональные блоки мозга Дифференциация систем мозговой коры происходит постепенно, и это приводит к неравномерному созреванию отдельных мозговых структур, ...
... . Кроме того, ДНК имеется в митохондриях, которые передаются только по материнской линии. Исследования мутаций в этих ДНК позволяют реконструировать историю биологического развития человечества, происхождение отдельных рас и народностей. Мутация может быть рецессивной, доминантной и полудоминантной в зависимости от состояния гена, в котором она произошла. Гены мутируют с определенной частотой, и ...
— концепция «2К» (кооперация и коммуникация) — эта концепция была разработана в 1900 г. русским ученым-философом П.А.Кропоткиным (1842—1921). В ней идет речь в основном о механизме биологической эволюции. . Термин «эволюция» (лат. evolutio — развертывание) в науке о живом представляет фундаментальное понятие для объяснения возникновения и развития всего живого. Эволюция подразумевает необратимое ...
... о противоречивом пути развития синтетической теории эволюции, о преодолении кризисов в научном познании, направленном на более полное и глубокое понимание закономерностей развития органического мира. Важно отметить, что основы научной теории эволюции заложил Ч. Дарвин. Как господствующее эволюционное учение дарвинизм существовал с 1859 до 1900 гг., т.е. до переоткрытия законов Г. Менделя. До ...
0 комментариев