Введение
Менее жестко запрограммированная эволюция
Выводы
Осложнения
Хромосомные и химические основы наследственности
Связь классической и молекулярной генетики
Неодарвинизм и популяционная генетика
Холдейн, Фишер и значение отбора
Сьюэлл Райт, адаптивные ландшафты и случай
Рекомендуемая литература
Меланизм у бабочек — подход a posteriori
Приложение теории игр к поведению животных
Изменчивость в природных популяциях; ее значение для адаптации
Адаптации на благо группы и адаптации групп
Адаптации самого полового размножения как такового и порожденные им адаптации
Рекомендуемая литература
Канализация развития
Заключения
Кладизм (5, 7)
Эволюционная систематика (13)
Как образуются таксоны — неодарвинистская точка зрения, или синтетическая теория эволюции
Альтернативная точка зрения — прерывистая эволюция
Сравнение теорий постепенной и прерывистой эволюции
Заключения
Навигация
Выводы
Принципы эволюции
242666
знаков
12
таблиц
37
изображений
1.6. Выводы.
В заключение следует еще раз подчеркнуть различие между а) точкой зрения о том, что жизнь возникла в процессе эволюции, и б) теориями о том, как происходила эволюция. К тому времени, когда было опубликовано «Происхождение видов», многие были готовы признать внушительный ряд данных, свидетельствующих в пользу эволюции, и в частности:
1) существование ископаемых остатков, свидетельствующих об изменениях фауны и флоры с течением времени;
2) сходство, лежащее в основе морфологии, физиологии и эмбриологии разных видов, наводящее на мысль об их общих предках;
3) географическая прерывистость в фаунах и флорах, связанная с физическими преградами (например, Австралия с ее своеобразными сумчатыми млекопитающими или острова — подобно Галапагосской группе, которые посетил и описал Дарвин,— с характерными для них одних видами растений и животных), заставляла предполагать естественный процесс изменения признаков у видов, оказавшихся «в ловушке» в этих изолированных областях.
Были возможны, однако, и многие другие объяснения эволюционного изменения. Дарвин выдвинул одно объяснение — естественный отбор случайных изменений, — менее антропоморфное, чем другие, и подтверждавшееся рядом наблюдений, в частности:
1) у особей, составляющих природные популяции, наблюдается значительная и притом часто непрерывная изменчивость;
2) путем отбора такой изменчивости животноводы добиваются внушительных морфологических и физиологических изменений у домашних животных;
3) адаптации основаны на модификации обычных принципов и зачастую бывают не так хороши, как могли бы быть, если бы они создавались de novo, без ограничений, налагаемых пред существующими структурами. Несовершенные решения, явно бесполезные признаки и рудиментарные органы, служат — несколько парадоксальным образом — доводом в пользу процесса эволюции, основанного на естественном отборе случайных изменений в рамках ограничений, создаваемых предсуществующими формами. Их трудно совместить с концепцией всемогущего творца.
1.7. Рекомендуемая литература.
Общая история биологии хорошо изложена в книге Смита (9). Полные обзоры истории, философии и социального значения дарвинизма содержатся в книгах Джиллеспи (2), Моора (4) и Рьюза (7, 8). Сведения о Дарвине как человеке и его путешествии на «Бигле» можно найти у Мурхеда (5), Джорджа (11 и Ховарда (3).
Глава 2. МЕХАНИЗМЫ НАСЛЕДСТВЕННОСТИ.
2.1. Механизм наследственности по Менделю.
2.1.1. Основные принципы.
Дарвин сознавал, что для правильной оценки эволюционных процессов очень важно отчетливо понимать явление наследственности. А между тем он завяз в теории, согласно которой наследственность работала против естественного отбора, «разбавляя», а не «концентрируя» благоприятные варианты. Человеком, разработавшим теорию наследственности, более совместимую с дарвиновской эволюцией, и заложившим основы современной генетики, был Грегор Мендель (Gregor Mendel, 1822—1884) В 1865 г. он сообщил о результатах своих экспериментов по гибридизации растений на заседании Общества естествоиспытателей в Брюнне (Брно), а на следующий год опубликовал их в «Трудах» этого общества. Однако его работа почти не привлекала внимания до тех пор, пока в 1900 г. Корренс (Correns) в Германии, де Фриз (de Vries) в Голландии и Чермак (Tschermak) в Австрии не открыли обнаруженные Менделем закономерности вторично и не оценили их значение и важность для теории Дарвина.
Изучая горох (Pisum sativutn), Мендель сумел выбрать у этого растения ряд особенностей, удобных для проведения экспериментов: 1) оно представлено многочисленными формами, четко отличающимися друг от друга по признакам, относящимся к семенам и к растению в целом; 2) предоставленные самим себе, эти растения могут самоопыляться, так как в каждом цветке имеются и тычинки, и пестики; 3) возможно проведение контрольных скрещиваний путем удаления тычинок из цветков (для предотвращения самоопыления) и опыления цветков пыльцой,, взятой от других растений.
Мендель выбрал для своей работы несколько признаков (табл. 2.1) и путем многократных самоопылений вывел чистые линии по этим признакам. Таким образом он установил, что растения размножаются по этим признакам в чистоте на протяжении многих поколений, из чего следовало, что контролирующие эти признаки наследственные факторы, по-видимому, очень стабильны. Проводя дальнейшие экспериментальные скрещивания, Мендель заметил, что при скрещивании двух чистых линий, различающихся по какой-либо паре признаков, все потомки (первое дочернее поколение, или f1) несут только один из этих признаков. Если, однако, скрещивать между собой растения этого первого поколения, то следующее поколение (поколение F2) содержит растения с признаками своих родителей, а также растения с исходными признаками, отсутствовавшими в поколении fi. Это означает, что признаки исходных растений выщепляются в F2, а следовательно, в fi не происходит слияния генетического материала. Наконец, Мендель обнаружил, что в поколении F2 признаки одного и другого типа появляются в определенном соотношении — примерно 3:1.
Таблица 2.1. Признаки, изучавшиеся Менделем (доминантные признаки набраны прописными.
| Признак | Варианты |
| Форма семян | ГЛАДКИЕ и морщинистые |
| Окраска семян | ЖЕЛТЫЕ и зеленые |
| Оболочка семени | ОКРАШЕННАЯ и белая |
| Бобы Окраска боба | ГЛАДКИЕ и морщинистые ЗЕЛЕНАЯ и желтая |
| Цветки | ПАЗУШНЫЕ и верхушечные |
| Стебель | ДЛИННЫЙ и короткий |
Мендель интерпретировал эти данные следующим образом: 1) наследственные детерминанты (он называл их факторами, а теперь мы называем их генами) дискретны; 2) они не сливаются; 3) у каждой взрослой особи имеется по два таких детерминанта (теперь их называют аллелями) для каждого признака, и они распределяются между гаметами — по одному на каждую гамету; 4) каждая взрослая особь может нести либо два идентичных аллеля (такую особь теперь называют гомозиготной), либо два разных аллеля (гетерозиготная особь), и в последнем случае экспрессия одного аллеля доминирует над экспрессией Другого (табл. 2.1); 5) гаметы соединяются случайным образом. Пункт 3 часто называют первым законом Менделя. Он означает, что гетерозиготные родительские особи производят в равных количествах гаметы, содержащие контрастирующие аллели.
Мендель обозначал пары генов буквами; доминантные аллели— прописными, а рецессивные — строчными. На рис. 2.1 эти обозначения использованы для наглядного изображения результатов одного из его экспериментов и их истолкования. Мендель скрещивал растения гороха с гладкими (доминантный признак— R) и морщинистыми (рецессивный признак — г) семенами и получил в поколении F2 5474 гладких и 1850 морщинистых семян, то есть отношение гладких семян к морщинистым составляло 2,96: 1, что совпадает с предсказанным соотношением 3:1.
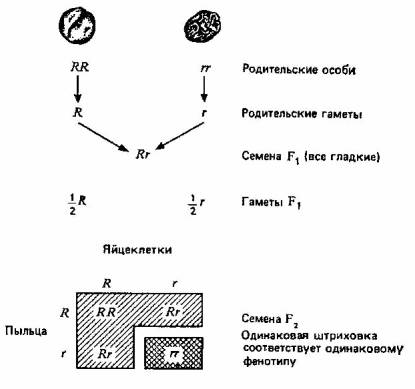
Предсказанное соотношение = 3 гладких :1 морщинистое
Рис. 2.1. Один из экспериментов Менделя по скрещиванию растений, различающихся по одной паре признаков.
Далее Мендель обнаружил, что если исходные растения, и с-! пользованные в скрещивании, различаются еще по двум контрастирующим признакам, то эти признаки в поколении fs также расщепляются, но при этом не обязательно остаются связанными с первой парой признаков. На основе этого Мендель сформулировал свой второй закон, или принцип независимого распределения: гены, детерминирующие разные признаки, распределяются по гаметам независимо друг от друга. (Последнее далеко не всегда справедливо, но это было установлено позднее.) Один из экспериментов Менделя по изучению наследования двух признаков и истолкование полученных данных представлены на рис. 2.2. Интересные результаты наблюдаются в поколении F2. Существует две возможности: 1) признаки, полученные от одной из родительских особей, передаются совместно, и в таком случае следует ожидать в поколении F2 семена только двух типов — гладкие-желтые и морщинистые-зеленые — в соотношении 3:1, как и прежде (рис. 2.1); 2) эти признаки передаются независимо, как показано на рис. 2.2. В последнем случае в fz должно быть 4 типа семян в соотношении 9 гладких-желтых (доминантный—доминантный), 3 гладких-зеленых (доминантный—рецессивный), 3 морщинистых-желтых (рецессивный—доминантный), 1 морщинистый-зеленый (рецессивный—рецессивный). Мендель получил семена этих четырех типов в соотношении 315 : 108 : 101 : 32, что хорошо соответствует пункту 2.
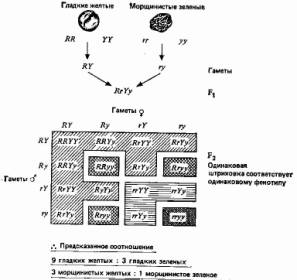
Рис. 2.2. Еще один эксперимент Менделя, в котором скрещивали растения, различающиеся по двум парам признаков.
Похожие работы
... матери требовали максимальной "притирки" плода к условиям организма матери, поэтому генетический контроль должен был быть минимальным. 2. Рост мозга. Развитие коры головного мозга. Развитие интеллекта. Эмоциональное развитие. 2.1 Функциональные блоки мозга Дифференциация систем мозговой коры происходит постепенно, и это приводит к неравномерному созреванию отдельных мозговых структур, ...
... . Кроме того, ДНК имеется в митохондриях, которые передаются только по материнской линии. Исследования мутаций в этих ДНК позволяют реконструировать историю биологического развития человечества, происхождение отдельных рас и народностей. Мутация может быть рецессивной, доминантной и полудоминантной в зависимости от состояния гена, в котором она произошла. Гены мутируют с определенной частотой, и ...
— концепция «2К» (кооперация и коммуникация) — эта концепция была разработана в 1900 г. русским ученым-философом П.А.Кропоткиным (1842—1921). В ней идет речь в основном о механизме биологической эволюции. . Термин «эволюция» (лат. evolutio — развертывание) в науке о живом представляет фундаментальное понятие для объяснения возникновения и развития всего живого. Эволюция подразумевает необратимое ...
... о противоречивом пути развития синтетической теории эволюции, о преодолении кризисов в научном познании, направленном на более полное и глубокое понимание закономерностей развития органического мира. Важно отметить, что основы научной теории эволюции заложил Ч. Дарвин. Как господствующее эволюционное учение дарвинизм существовал с 1859 до 1900 гг., т.е. до переоткрытия законов Г. Менделя. До ...
0 комментариев