Введение
Менее жестко запрограммированная эволюция
Выводы
Осложнения
Хромосомные и химические основы наследственности
Связь классической и молекулярной генетики
Неодарвинизм и популяционная генетика
Холдейн, Фишер и значение отбора
Сьюэлл Райт, адаптивные ландшафты и случай
Рекомендуемая литература
Меланизм у бабочек — подход a posteriori
Приложение теории игр к поведению животных
Изменчивость в природных популяциях; ее значение для адаптации
Адаптации на благо группы и адаптации групп
Адаптации самого полового размножения как такового и порожденные им адаптации
Рекомендуемая литература
Канализация развития
Заключения
Кладизм (5, 7)
Эволюционная систематика (13)
Как образуются таксоны — неодарвинистская точка зрения, или синтетическая теория эволюции
Альтернативная точка зрения — прерывистая эволюция
Сравнение теорий постепенной и прерывистой эволюции
Заключения
Навигация
Адаптации на благо группы и адаптации групп
Принципы эволюции
242666
знаков
12
таблиц
37
изображений
3.6. Адаптации на благо группы и адаптации групп.
До сих пор теория неодарвинизма развивалась на основе допущения, что частота того или иного гена возрастает, если он детерминирует признак, который повышает выживаемость и (или) репродукцию его обладателя. Между тем у животных можно найти много примеров такого поведения, которое, очевидно, не способствует выживанию проявляющих его особей. Например, у общественных насекомых некоторые касты лишены способности к размножению, с тем чтобы они могли более эффективно участвовать в различных работах в улье. Рабочая пчела защищает свой улей, убивая вторгшегося в него чужака, даже если погибает при этом сама. Как могло возникнуть в процессе эволюции такое альтруистическое поведение?
Решить этот вопрос удалось после того, как в наших представлениях о приспособленности был достигнут существенный прогресс благодаря работам У. Д. Гамильтона (19). Его рассуждения основаны на том, что родственные особи несут сходные гены, причем сходство между генами тем больше, чем теснее родство. Поэтому если какой-либо признак может повысить выживаемость других особей, обладающих этим же самым признаком, то это может привести к повышению частоты гена, детерминирующего этот признак, даже если данный носитель признака при этом погибнет. Проявления подобного рода альтруизма, вероятно, выше между особями, связанными близким родством, и соответствующий процесс получил название К-отбора (от англ, kin — родство).
Вклад Гамильтона (19) в развитие этой проблемы состоял в том, что он формализовал эти рассуждения и сделал их более строгими. Допустим, что некий альтруистический признак детерминируется одной парой аллелей, А к а, причем аллель А повышает вероятность альтруистического акта у несущей его особи, например делает более вероятным, что эта особь, рискуя собственной жизнью, будет спасать молодых особей, отвлекая на себя внимание хищника. Совершая этот акт, взрослая особь снижает свои шансы на выживание на величину С и повышает соответствующие шансы молодых особей на величину В. Гамильтон показал, что частота в популяции аллеля А по отношению к аллелю а будет возрастать только в том случае, если родство (rl) между донором и реципиентом (выигрывающим от альтруистического акта) достаточно близкое; выражаясь точнее, rl должно быть больше чем С/В.
Например, в случае родители—потомки вероятность того, что любой из генов, имеющихся у одной из родительских особей, окажется у ее потомка, равна 0,5, поскольку потомок наследует гены от обеих родительских особей. Следовательно, rl= = 0,5, и в соответствии с утверждением Гамильтона альтруистический признак сможет спасти более чем двух потомков, прежде чем частота аллеля А повысится относительно частоты аллеля а. В действительности величины rl выражают вероятность того, что организмы, связанные друг с другом определенными родственными отношениями, будут нести одни и те же гены, а тем самым — вероятность того, что реципиент получит гены, определяющие альтруистический признак (несколько других примеров приведено в табл. 3.4). При альтруистических взаимоотношениях между соответствующими родичами с уменьшением rl действия донора должны приносить пользу все большему и большему числу реципиентов; иначе гены альтруизма распространяться не будут. Следовательно, вероятность сотрудничества выше между близкородственными особями.
Идеи Гамильтона помогают понять даже некоторые аспекты организации сообщества у перепончатокрылых. У этих насекомых самки развиваются из оплодотворенных яиц и поэтому
Таблица 3.4. Коэффициенты родства (rl) между особями при разной степени родства.
| Родител и / потомки | 0,5 |
| Бабки-деды /вн уки | 0,25 |
| Полные сибсы | 0,5 |
| Полусибсы | 0,25 |
| Дядя/племянник | 0,125 |
| Двоюродные сибсы | 0,125 |
диплоидны, но самцы развиваются из неоплодотворенных яиц и поэтому гашгоидны. В популяции, в которой особи обоих полов диплоидны, коэффициент родства между сибсами такой же, как между одной из родительских особей и ее потомком (г/ = 0,5; см. табл. 3.4). Однако у самки перепончатокрылых в результате гапло-диплоидии больше общих генов с родной сестрой, чем с собственными дочерьми; то есть генотипы сестер идентичны по всем генам, которые они получают от своих отцов (у него всего один набор), и в среднем по половине генов, которые они получают от своих матерей. Поэтому коэффициент родства между родными сестрами у перепончатокрылых равен не 0,5; а (0,5) X Х(1) + (0,5) (0,5)— 0,75. Таким образом, в этом случае дочерям следует помогать своим матерям охранять яйца и снабжать кормом молодь, а не производить собственных потомков, так что генетическая композиция перепончатокрылых предрасполагает их к развитию такого сообщества, в котором стерильные рабочие самки заботятся о своих родных сибсах. Из всего этого вытекают и дальнейшие следствия, для ознакомления с которыми мы отсылаем читателя к работе Гамильтона (20). Сравнительно недавно идеи Гамильтона были использованы в изучении общественной жизни высших организмов — птиц, млекопитающих и даже человека (см. (44); критические замечания см. (35)).
К-отбор не может автоматически вести к закреплению признаков, полезных для данного вида (то есть обеспечивающих ему длительное существование; см. разд. 2.4); он ведет лишь к признакам, полезным для родичей. Тем не менее можно представить себе способы эволюции таких признаков «на благо вида» путем отбора самих групп, а не особей внутри этих групп. Представим себе две полуизолированные популяции какого-либо вида хищников. В одной из них возникает мутация, порождающая «сверххищника», который обнаруживает и схватывает жертву более эффективно, чем другие особи. Частота такого мутанта в популяции должна возрастать, несмотря на то, что это приведет к вымиранию жертвы, а в конечном счете и самого хищника. В другой популяции такая мутация не возникла, так что она продолжает существовать и в конце концов может захватить пространство, которое занимала вымершая популяция. Аналогичный результат получится в том случае, если от популяции «сверххищников» отделится субпопуляция, не содержащая гена «сверххищника», например, вследствие эффекта основателя.
Можно привести и несколько иной пример: представим себе снова две субпопуляции, но состоящие на этот раз из бессмертных особей. В одной из них появляется мутация, вызывающая старение, и это оказывается выгодным для данной группы, потому что ограничивает ее численность, «очищает» от изношенных особей и увеличивает простор для благоприятных мутаций. Группа, в которой возникло старение, сохраняется дольше другой группы. Этот пример отличается от примера со сверххищниками, потому что он связан с эволюцией признака, положительного для группы, — старения, а не отрицательного — «быть сверххищником». Труднее представить себе, как это может реализоваться. Как, например, ген старения может закрепиться в субпопуляции? Для закрепления такого гена необходимы либо повторные мутации, либо дрейф, либо эффект основателя, поскольку он не может закрепиться при помощи отбора: бессмертные особи оставляют больше потомков, чем смертные, так что ген, обусловливающий старение, будет элиминироваться.
Какие существуют данные за и против группового отбора?
1. Возможен ли групповой отбор? Для того чтобы групповой отбор был реально возможен, скорость вымирания групп и (или) скорость образования новых групп (не имеющих эгоистичного гена или имеющих альтруистический ген) должна быть выше скорости поступления эгоистичных генов в результате либо мутационного процесса, либо, что более важно, обмена генами между подгруппами. По мнению одних биологов, это слишком жесткое требование, чтобы групповой отбор мог быть обычным явлением (например, (24), тогда как другие этого не считают (например, (17, 43)).
2. Происходит ли групповой отбор? Располагаем ли мы данными о существовании признаков, важных для сохранения группы, но нестабильных при индивидуальном отборе? Один из таких возможных признаков — это старение, и оно широко распространено. Однако его можно объяснить также, как плейот-ропный эффект благоприятных генов, которые отбирались обычным образом (см. разд. 3.2). Другой возможный пример — способность популяций регулировать свою численность, поддерживая ее на таком уровне, чтобы не допускать чрезмерного изъятия ресурсов. Винн-Эдвардс в своей знаменитой книге «Расселение животных и его зависимость от социального поведения» (Wynne-Edwards, Animal Dispersion in Relation to Social Behaviour, Oliver a. Boyd, 1962) привел многочисленные примеры такого рода явлений. Однако при более тщательном изучении то, что представляется такой саморегуляцией, на самом деле часто можно объяснить как результат зависящей и не зависящей от плотности регуляции, обусловленной внешними факторами — хищниками, погодой и даже самими ресурсами. Многие другие групповые признаки оказалось возможным объяснить К-отбором.
Данные, свидетельствующие за и против группового отбора, неубедительны, и поэтому данная проблема продолжает вызывать разногласия. Представляется вероятным, что в некоторых системах может происходить и действительно происходит групповой отбор, но вопрос о его относительной частоте по сравнению с индивидуальным отбором остается открытым. Возможно также, что существует групповой отбор сходного типа, действующий, однако, не во внутривидовых группах, а на уровне вида в целом (см. гл. 5).
Похожие работы
... матери требовали максимальной "притирки" плода к условиям организма матери, поэтому генетический контроль должен был быть минимальным. 2. Рост мозга. Развитие коры головного мозга. Развитие интеллекта. Эмоциональное развитие. 2.1 Функциональные блоки мозга Дифференциация систем мозговой коры происходит постепенно, и это приводит к неравномерному созреванию отдельных мозговых структур, ...
... . Кроме того, ДНК имеется в митохондриях, которые передаются только по материнской линии. Исследования мутаций в этих ДНК позволяют реконструировать историю биологического развития человечества, происхождение отдельных рас и народностей. Мутация может быть рецессивной, доминантной и полудоминантной в зависимости от состояния гена, в котором она произошла. Гены мутируют с определенной частотой, и ...
— концепция «2К» (кооперация и коммуникация) — эта концепция была разработана в 1900 г. русским ученым-философом П.А.Кропоткиным (1842—1921). В ней идет речь в основном о механизме биологической эволюции. . Термин «эволюция» (лат. evolutio — развертывание) в науке о живом представляет фундаментальное понятие для объяснения возникновения и развития всего живого. Эволюция подразумевает необратимое ...
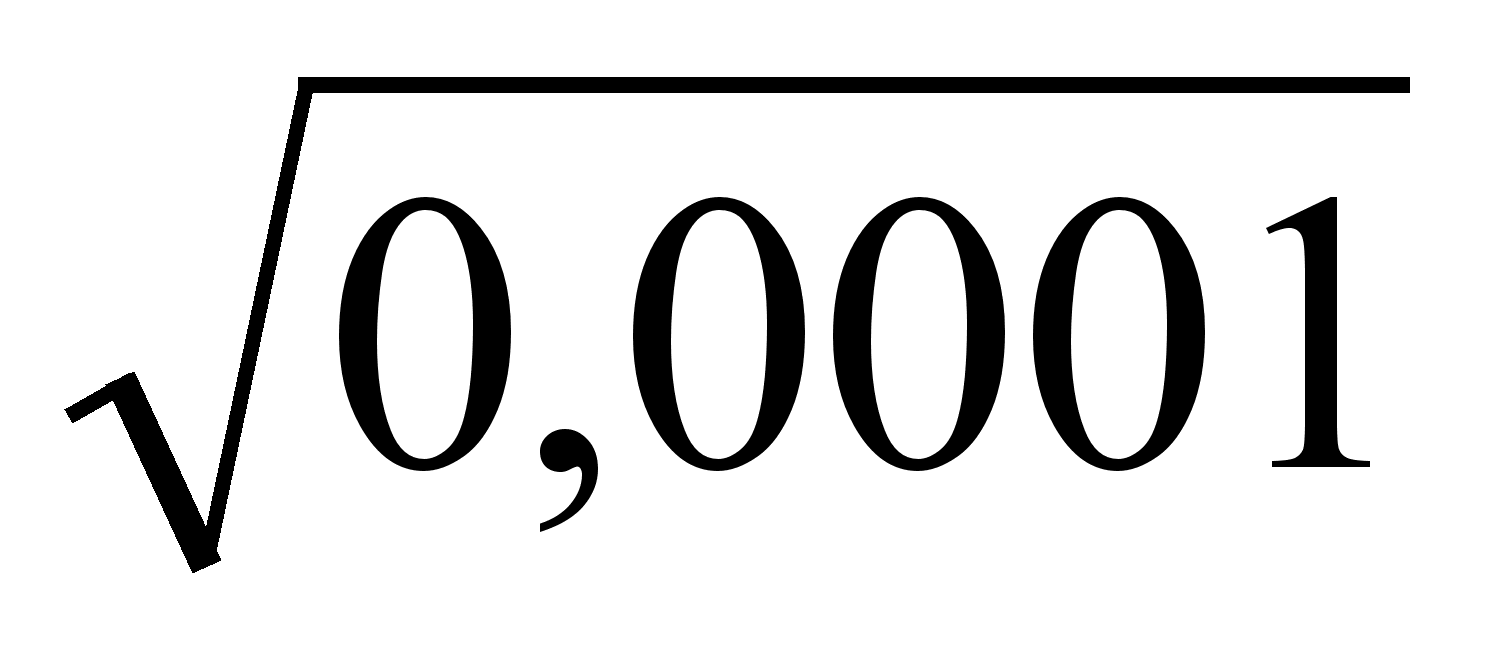
... о противоречивом пути развития синтетической теории эволюции, о преодолении кризисов в научном познании, направленном на более полное и глубокое понимание закономерностей развития органического мира. Важно отметить, что основы научной теории эволюции заложил Ч. Дарвин. Как господствующее эволюционное учение дарвинизм существовал с 1859 до 1900 гг., т.е. до переоткрытия законов Г. Менделя. До ...
0 комментариев