Введение
Менее жестко запрограммированная эволюция
Выводы
Осложнения
Хромосомные и химические основы наследственности
Связь классической и молекулярной генетики
Неодарвинизм и популяционная генетика
Холдейн, Фишер и значение отбора
Сьюэлл Райт, адаптивные ландшафты и случай
Рекомендуемая литература
Меланизм у бабочек — подход a posteriori
Приложение теории игр к поведению животных
Изменчивость в природных популяциях; ее значение для адаптации
Адаптации на благо группы и адаптации групп
Адаптации самого полового размножения как такового и порожденные им адаптации
Рекомендуемая литература
Канализация развития
Заключения
Кладизм (5, 7)
Эволюционная систематика (13)
Как образуются таксоны — неодарвинистская точка зрения, или синтетическая теория эволюции
Альтернативная точка зрения — прерывистая эволюция
Сравнение теорий постепенной и прерывистой эволюции
Заключения
Навигация
Сьюэлл Райт, адаптивные ландшафты и случай
Принципы эволюции
242666
знаков
12
таблиц
37
изображений
2.3.3. Сьюэлл Райт, адаптивные ландшафты и случай.
Приведенные выше анализы были основаны на рассмотрении пар аллелей, находящихся в одном и том же локусе. Однако большая часть фенотипических признаков контролируется совокупностями генов с участием многих аллелей и локусов. Эти гены могут быть в разной степени сцеплены друг с другом; кроме того, между ними могут происходить различные взаимодействия (см. разд. 2.1.2). Следовательно, приспособленность определенных аллелей в одном локусе зависит от аллелей, лежащих в дру* гих локусах, и, в сущности, на нее оказывает влияние вся генетическая среда, в которой они находятся. Быть может, группы сцепления, целые хромосомы или даже геномы представляют собой более значимые единицы отбора, чем отдельные гены (см. разд. 3.8). К сожалению, генетика таких полигенных систем чрезвычайно сложна, а поэтому мы не можем охватить ее в такой мере, как случай двух аллелей, лежащих в одном локусе. Мы исходим из допущения, что основные принципы остаются теми же, даже если существуют различия в деталях. Однако здесь возможны серьезные исключения.
Другой основатель неодарвинизма, Сьюэлл Райт (Sewall Wright), разработал концептуальную систему, позволяющую рассмотреть взаимодействие между генами и их влияние на приспособленность (см. Wright, 1931, Genetics, 16, 99—159). Она получила название адаптивного ландшафта или пространства приспособленности. В этой системе каждая из осей я-мерного пространства определяет частоту каждого гена в данной попу-ляции,_причем последняя ось определяет среднюю приспособленность W, так что каждое сочетание генов определяет некую частную приспособленность. Так, в простейшей из возможных трехмерной модели две оси соответствуют частоте двух аллелей в каждом из двух локусов, а пространство между этими осями — всем возможным сочетаниям этих генов. Третья ось соответствует значениям W для каждого сочетания. Итак, мы имеем трехмерный ландшафт (более сложные ситуации порождают п-мер-ный ландшафт, с которым столь же трудно иметь дело, как и просто представить его себе). Пример адаптивного ландшафта изображен на рис. 2.8. В этой модели наиболее приспособленная популяция находится на вершине «холмов», а действие отбора направлено на то, чтобы заставить популяции подниматься на вершины холмов и закрепляться на них. Не все адаптивные пики будут или должны быть заняты, поскольку число потенциально возможных сочетаний аллелей у одного вида больше числа самих видов. В связи с этим многие пики ни разу не будут «опробованы» и некоторые из них будут выше чем другие. На какой пик поднимутся популяции, зависит от близости популяций к этим пикам и, следовательно, от исходных генетических условий до начала отбора. В этом теория Райта существенно отличается от модели «два аллеля — один локус», которая предсказывает, что W всегда будет максимальным (то есть популяции должны подниматься только на самые высокие пики), а из того обстоятельства, что это не есть непременное условие, вытекает ряд следствий, имеющих значение для вопросов, относящихся к адаптациям организмов, которые мы рассмотрим в гл. 3.
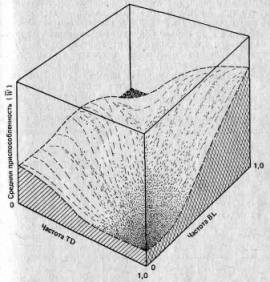
Рис. 2.8. Адаптивный ландшафт при различных частотах двух хромосомных инверсий у одного из представителей прямокрылых. BL — инверсия Blundell; TD —инверсия Tidbinbilla. (Shorrocks В., The Genesis of Diversity, Hodder a. Stoughton, London, 1978, по данным Lewontin R. C., White M. J. D., Evolution, 1960, 13, 561—564.)
На рис. 2.9 адаптивные ландшафты использованы для того, чтобы представить в качественном аспекте некоторые возможные эволюционные ситуации. (Следует помнить, что мы рассматриваем эти ландшафты сверху, то есть видим их контуры.) На рис. 2.9, А показано, чего следует ожидать в случае ослабления действия отбора или усиления мутационного процесса: область, занимаемая некой популяцией, распространяется по склонам ее адаптивного пика, и если это происходит достаточно интенсивно, то вид, к которому она принадлежит, будет вынужден занять еще один пик. На рис. 2.9, Б изображены результаты усиления отбора; в этом случае популяция оказывается сосредоточенной на высшем уровне своего адаптивного пика. Рис. 2.9, В иллюстрирует результаты изменений в среде; в этом случае рельеф ландшафта изменяется, так что адаптивный пик может «рухнуть» под тяжестью популяции, которая таким образом окажется в «долине». Затем под действием отбора популяция начнет подниматься на какой-либо близлежащий пик.
Как показывают рис. 2.9, Л и Б, для того чтобы популяция оставалась на занятом ею пике, необходим непрерывный отбор; такой отбор получил название стабилизирующего отбора: в результате, непрерывного отбора против мутантов популяция сохраняет стабильность в отношении как морфологических, так и физиологических признаков. Процесс В, хотя в его основе лежат те же механизмы, приводит к другим результатам: под его воздействием популяция приобретает новое адаптивное сочетание признаков. Такой отбор получил название направленного отбора.
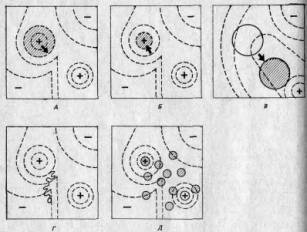
Рис. 2.9. Несколько адаптивных ландшафтов — вид сверху. А. Ослабление отбора или усиление мутационного процесса. Б. Стабилизирующий отбор. В. Направленный отбор. Г. Случайный дрейф-Д. Популяция, разделившаяся на демы. (Wadd-ington С. Н., An Introduction to Modern Genetics, George Alien a. Unvin Ltd, London, 1939.)
На рис. 2.9, Г представлена небольшая популяция. Здесь случайные события могут вызвать сдвиг в сторону от адаптивного пика, несмотря на отбор, и это та возможность, которую Сьюэлл Райт сформулировал вполне определенно. В принципе каждый индивидуум может случайно стать жертвой несчастного случая или же не найти себе брачного партнера, в результате чего он не внесет своего вклада в следующее поколение независимо от того, сколь хорошо он приспособлен. Поэтому гены, которые он несет, как бы хороши они ни были потенциально, не будут представлены в следующем поколении. Для большой популяции это не будет иметь существенного значения. Однако в небольших популяциях такие случайные события могут оказать серьезное воздействие на частоты генов, так как эти частоты определяются небольшим числом особей. При таких обстоятельствах популяция может медленно отойти от адаптивного пика; процессе этот получил название генетического дрейфа. Подобным же образом при резком сокращении численности популяции или же, что, в сущности, то же самое, при образовании несколькими особями-основателями новой популяции выборка генов из генофонда исходной популяции может произойти неслучайным образом. В результате будет иметь место некое начальное изменение, не связанное с отбором. Это явление известно под названием эффекта основателя.
На рис. 2.9, Д изображена большая популяция, разделенная на ряд субпопуляций, каждая из которых занимает части данного адаптивного ландшафта, несколько отличающиеся друг от друга. Такие популяции, вероятно, встречаются довольно часто. Степень отклонения каждой группы (в результате отбора или дрейфа) от среднего генетического состава популяции в целом зависит от интенсивности обмена генами (то есть миграции индивидуумов) между группами. Для того чтобы противодействовать отбору, поступление данного гена в субпопуляцию должно превышать его элиминацию отбором, а для того, чтобы противодействовать дрейфу, — превышать случайные потери этого гена. Когда такие субпопуляции более или менее изолированы, их называют демами. Демовая структура потенциально важна для эволюционных процессов, и мы обсудим этот вопрос в разд. 3.6. Генетический дрейф иногда называют эффектом Сьюэлла Райта и считают, что его действие направлено против действия отбора (рис. 2.9,Г). Однако, как это понимал Сьюэлл Райт, генетический дрейф может дать популяциям возможность покинуть локальные пики, пересечь имеющиеся в адаптивном ландшафте долины и подпасть под влияние факторов отбора, связанных с новыми пиками. Следовательно, генетический дрейф может способствовать адаптации, но может и препятствовать ей.
2.4. Выводы и резюме.
В процессе исторического развития генетических основ эволюции мы были свидетелями ряда важных переключений внимания с одних факторов на другие.
1. Сторонники Менделя уделяли главное внимание последствиям скрещивания пар особей с известной генетической конституцией. Их основное допущение заключалось в том, что признаки контролируются дискретными факторами, которые не сливаются друг с другом.
2. Холдейн и Фишер рассматривали эволюционное изменение, исходя из частот отдельных генов в популяциях.
3. Для Райта, однако, главными были популяции генов (то есть величины W, а не w), а Добржанский предложил обозначать их термином «генофонд» (см. Th. Dobzhansky, Genetics and the Origin of Species, Columbia Univ, Press, 1937).
4. Начиная с работ Вейсмана, ученые постепенно приближались к открытию физической основы наследственности. В конечном итоге это привело к переосмыслению концепций генов,, мутаций и т. п. в свете представлений о ДНК и синтезе белков.
Первая из этих позиций соответствует менделевской генетике, вторая и третья — популяционной генетике, а четвертая — молекулярной генетике. Позиции 2 — 4 составляют неодарвинизм.
Здесь уместно еще раз привести взгляды, которых придерживается большинство неодарвинистов. (Не все неодарвинисты разделяют в точности одни и те же взгляды, и различия в расстановке акцентов не раз оказывались причиной возникавших & дальнейшем разногласий.) Главные допущения неодарвинизма состоят в следующем.
1. В основе эволюции лежат генетические изменения.
2. Мутации (в их современном общем смысле) представляют собой в конечном счете источник изменчивости. Это могут быть мелкие (точковые) или крупные (хромосомные) мутации, однако мелкие мутации скорее могут привести к благоприятным изменениям, чем крупные, а поэтому они играют более важную роль в эволюции. Принципиально важное значение имеет тот факт, что эти мутации случайны, иными словами, они не направленны.
3. Центральная догма и принцип Вейсмана принимаются.
4. Эволюция осуществляется путем изменения частот генов.
5. Эти изменения могут происходить в результате мутаций, поступления генов в популяцию и оттока их из нее, случайного дрейфа и естественного отбора, причем отбору принадлежит наиболее важная роль.
6. Перечисленных принципов достаточно, для того чтобы объяснить разнообразие и адаптацию организмов на земном шаре.
Осталось еще несколько вопросов, к которым мы вернемся в дальнейших разделах. Это, во-первых, вопрос об • относительном участии генотипа и среды в формировании фенотипа (см. разд 3.2 и гл. 4). Во-вторых, кратковременные изменения генных частот в популяциях мало говорят нам о сохранении самих этих популяций в течение длительного времени. Между тем мы интуитивно ощущаем, что это последнее представляет собой по меньшей мере столь же важную меру эволюционного успеха, как и первое. Можно ли определить это долговременное сохранение с позиций неодарвинизма? И совместимо ли оно вообще с неодарвинизмом? Эти вопросы будут рассмотрены в разд. 3.6 и 5.4.
Похожие работы
... матери требовали максимальной "притирки" плода к условиям организма матери, поэтому генетический контроль должен был быть минимальным. 2. Рост мозга. Развитие коры головного мозга. Развитие интеллекта. Эмоциональное развитие. 2.1 Функциональные блоки мозга Дифференциация систем мозговой коры происходит постепенно, и это приводит к неравномерному созреванию отдельных мозговых структур, ...
... . Кроме того, ДНК имеется в митохондриях, которые передаются только по материнской линии. Исследования мутаций в этих ДНК позволяют реконструировать историю биологического развития человечества, происхождение отдельных рас и народностей. Мутация может быть рецессивной, доминантной и полудоминантной в зависимости от состояния гена, в котором она произошла. Гены мутируют с определенной частотой, и ...
— концепция «2К» (кооперация и коммуникация) — эта концепция была разработана в 1900 г. русским ученым-философом П.А.Кропоткиным (1842—1921). В ней идет речь в основном о механизме биологической эволюции. . Термин «эволюция» (лат. evolutio — развертывание) в науке о живом представляет фундаментальное понятие для объяснения возникновения и развития всего живого. Эволюция подразумевает необратимое ...
... о противоречивом пути развития синтетической теории эволюции, о преодолении кризисов в научном познании, направленном на более полное и глубокое понимание закономерностей развития органического мира. Важно отметить, что основы научной теории эволюции заложил Ч. Дарвин. Как господствующее эволюционное учение дарвинизм существовал с 1859 до 1900 гг., т.е. до переоткрытия законов Г. Менделя. До ...
0 комментариев