Введение
Менее жестко запрограммированная эволюция
Выводы
Осложнения
Хромосомные и химические основы наследственности
Связь классической и молекулярной генетики
Неодарвинизм и популяционная генетика
Холдейн, Фишер и значение отбора
Сьюэлл Райт, адаптивные ландшафты и случай
Рекомендуемая литература
Меланизм у бабочек — подход a posteriori
Приложение теории игр к поведению животных
Изменчивость в природных популяциях; ее значение для адаптации
Адаптации на благо группы и адаптации групп
Адаптации самого полового размножения как такового и порожденные им адаптации
Рекомендуемая литература
Канализация развития
Заключения
Кладизм (5, 7)
Эволюционная систематика (13)
Как образуются таксоны — неодарвинистская точка зрения, или синтетическая теория эволюции
Альтернативная точка зрения — прерывистая эволюция
Сравнение теорий постепенной и прерывистой эволюции
Заключения
Навигация
Меланизм у бабочек — подход a posteriori
Принципы эволюции
242666
знаков
12
таблиц
37
изображений
3.3. Меланизм у бабочек — подход a posteriori.
По мере развития промышленности в Великобритании энтомологи стали замечать, что у многих различных видов бабочек светлая окраска заменяется темной или черной. Особенно ярким примером этого служит березовая пяденица (Biston betularia), темная форма которой получила название carbonaria. Были обнаружены следующие факты: 1) самые ранние находки темных форм были сделаны вблизи районов с сильно развитой промышленностью; 2) самая высокая частота темных форм обнаружена вблизи промышленных центров (отсюда и название этого явления — индустриальный меланизм); 3) меланисты обычно встречаются у видов, которые активны ночью. Апостериорное эволюционное объяснение состоит в том, что светлые формы более заметны для хищников на закопченных ветвях и стволах деревьев, на которых они отдыхают днем. Существуют, конечно, и другие возможные объяснения. Прежде всего меланизм может иметь фенотипическую природу: например, он мог бы возникнуть в результате попадания в организм насекомых промышленных загрязнений. Однако в экспериментах по скрещиванию было показано, что меланизм контролируется генетически и что расщепление по этому признаку происходит так, как если бы он определялся одним доминантным аллелем. Есть и другие возможные объяснения. Так, например, темная окраска могла бы служить защитой от прямого действия загрязнения. Для дальнейшего изучения меланизма у березовой пяденицы Кеттлуэлл (21, 22) провел следующие полевые эксперименты.
1. Чтобы определить, не обладает ли форма carbonaria болы шей приспособленностью, чем типичная березовая пяденица, Кеттлуэлл выпускал меченых особей той и другой формы в промышленных и сельских районах. При повторном отлове в промышленных районах среди выловленных меченых особей было больше carbonaria, а в сельских — больше типичной формы.
2. Равное число особей carbonaria и типичной формы помещали на стволы деревьев в задымленной и незадымленной местностях и непосредственно наблюдали за тем, как их выедали хищники. Оказалось, что в сельской местности хищники уничтожили больше меланистических форм, а в промышленном районе— больше типичных особей.
3. Одна птица — пищуха — поедала без разбору обе формы. Кеттлуэлл указывает, что она кормится, перемещаясь вверх и вниз по стволам деревьев, и поэтому видит силуэт бабочки, сидящей на коре дерева, а не цвет ее крыльев.
Результаты экспериментов 2 и 3 служат особенно убедительными доводами в пользу того, что меланизм возник в ответ на давление со стороны хищников.
Приведенный здесь анализ индустриального меланизма основан на описанном выше методе 1, однако благодаря тщательно проведенным наблюдениям и экспериментам он зашел значительно дальше, чем это обычно бывает или чем это возможно. Кроме того, в этом случае удалось выявить генетическую основу наблюдаемого изменения. Подобного рода программы, основанные на методе 1, но учитывающие также экологические и генетические наблюдения, часто называют экогенетическими — подход, впервые примененный Е. Б. Фордом .(см. (16)) и Добр-Жанским (14).
3.4. Примеры априорного подхода.
3.4.1. Применение принципа оптимальности к адаптациям жизненного цикла
Большая часть рассматривавшихся до сих пор адаптации касалась либо морфологических, либо физиологических признаков. Между тем такие признаки, как характер морфогенетических процессов, скорости роста, общие размеры, продолжительность жизни и вклад в размножение, также, вероятно, подвергались естественному отбору. Все эти признаки участвуют в том, чтобы довести продукты размножения до такого состояния, в котором они могут воспроизводиться, и их называют признаками жизненного цикла или цикла развития. Со времени первой работы Лей-монта Коула (9) об эволюции жизненного цикла писали много (хорошие обзоры принадлежат Стирнсу (37, 38)).
Здесь в качестве иллюстрации мы рассмотрим количество ресурсов, затрачиваемых родительской особью на продуцирование гамет, и продолжительность пострепродуктивного периода ее жизни. То обстоятельство, что не все организмы бурно размножаются один раз в жизни, заставляет думать, что размножение сопряжено с большими затратами ресурсов. В противном случае приспособленность в неодарвинистском смысле была бы всегда максимизирована путем максимально возможного репродуктивного выхода, а следовательно, путем затраты максимального количества ресурсов на образование гамет. Принято считать, что, чем больше ресурсов данная родительская особь вкладывает в размножение, тем меньше их у нее остается для того, чтобы обеспечить защиту собственного организма от несчастных случаев, болезней и хищников, так что эти расходы можно оценивать в терминах выживаемости родительских особей. В результате с увеличением вклада в репродукцию шансы родителей на выживание в пострепродуктивный период снижаются; здесь мы используем фишеровскую меру приспособленности, то есть г (см. разд. 2.3.2), в сочетании с упрощенным вариантом принципа оптимальности, с тем чтобы исследовать, какого рода компромисс возможен между этими переменными.
Значение приспособленности г можно записать как
![]()
где It — выживаемость с момента рождения до времени t, a nt — число потомков, произведенных ко времени t (простой вывод этого уравнения см. у Уилсона и Боссерта (45)). В простейшем случае, когда размножение происходит один раз в год (t=1), плодовитость (п) зависит от возраста и родительские особи выживают после размножения, уравнение 3.1 принимает вид
![]()
где 1а и // — соответственно выживаемость взрослых особей и потомков, а п— число потомков на одну родительскую особь. Отсюда
![]()
Таким образом, из уравнения 6.6 мы видим, что если построить график зависимости 1а от п, то изоклины равных г будут иметь вид прямых с наклоном, равным — l1 (рис. 3.3.).
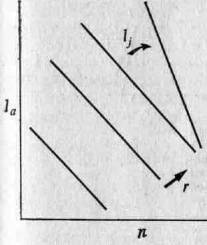
Рис. 3.3. На графике зависимости 1а от п изоклины т представляют собой прямые линии с отрицательными наклонами //. По мере увеличения выживаемости потомков наклон возрастает. В принципе приспособленность, то есть г, возрастает с увеличением la и п.
Однако вследствие компромисса между выживаемостью родительских особей и размножением не все комбинации 1а с п реально осуществимы. Можно представить себе некоторое число распределений реально возможных величин; одно из них показано на рис. 3.4. Здесь выживаемость взрослых особей снижается все быстрее, по мере того как все больше ресурсов вкладывается в размножение, то есть производится все больше гамет.
С помощью этих распределений нетрудно найти оптимальное решение: это та реально осуществимая комбинация 1а и п, которая лежит на самой высокой r-изоклине, то есть максимизирует г, или приспособленность. Для «компромиссной кривой», изображенной на рис. 3.4, этот оптимум приходится на крайнюю правую часть кривой; в этом случае отбор благоприятствует высокому уровню репродукции за счет родительской особи. Такой тип размножения называют семелопарией — родительская особь дает потомство один раз, после чего гибнет подобно лососю.
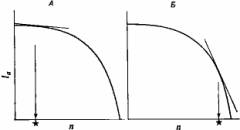
Рис. 3.4. Возможная форма истинной зависимости между 1а и п. А. Выживаемость потомков (//) низкая, "и поэтому отбор благоприятствует небольшому п (оптимум помечен звездочкой) при продолжительном выживании взрослых особей (высокое 1а). Значение // высокое, и поэтому отбор благоприятствует высокому п при низком 1а. Ситуация А ведет к итеропарии, а ситуация Б — к семелопарии (см. текст).
При альтернативном типе размножения, когда достигается некий компромисс между п и 1а, отбор благоприятствует многократному размножению, или итеропарии.
Перейдем теперь к предсказанию. С уменьшением шансов потомков на выживание (то есть с уменьшением /у) наклон г-изо-клин уменьшается и отбор благоприятствует итеропарии. При возрастании // справедливо обратное. (Точный результат зависит от формы «компромиссной» кривой, однако мы продолжаем допускать, что применима только кривая, изображенная на рис. 3.4.) Для того чтобы проверить это предсказание, нам необходимо найти популяции либо того же самого, либо других, но близкородственных видов с различными значениями //. На самом деле точных данных о возрастной специфичности выживаемости для популяций, находящихся в природных условиях, удивительно мало, однако имеются некоторые более или менее эпизодические данные, в общем подтверждающие эти предсказания. Например, пресноводные планарии (трехветвистокишечные тур-беллярии), вылупляясь из яиц, в природе попадают в плохие трофические условия, и, как показали наблюдения (6), молодые особи семелопарических видов гибнут не так быстро, как молодь итеропарических видов, голодающих в лабораторных условиях. Блауэр (3) описал семелопарические и итеропарические виды многоножек фауны Британских островов: число потомков на одну родительскую особь у первых было больше, чем у вторых. Семелопарические виды питаются листьями, которые равномерно покрывают землю в лесу, тогда как итеропарические виды более специализированы, и по крайней мере один из них питается и откладывает яйца на поваленных деревьях, которые разбросаны по лесу неравномерно. Расселение молоди по этим пятнисто распределенным ресурсам носит случайный характер, а поэтому значение // у этого вида ниже, чем у видов, питающихся листьями. Вероятно, по тем же причинам итеропарические виды с непрерывным или полунепрерывным размножением возникли у эндопаразитов, но в этом случае, поскольку родительская особь окружена сверхобильными пищевыми ресурсами — в виде тканей своего хозяина, — она может производить огромное число потомков и не должна расплачиваться за это сокращением продолжительности жизни, о котором говорилось выше.
Похожие работы
... матери требовали максимальной "притирки" плода к условиям организма матери, поэтому генетический контроль должен был быть минимальным. 2. Рост мозга. Развитие коры головного мозга. Развитие интеллекта. Эмоциональное развитие. 2.1 Функциональные блоки мозга Дифференциация систем мозговой коры происходит постепенно, и это приводит к неравномерному созреванию отдельных мозговых структур, ...
... . Кроме того, ДНК имеется в митохондриях, которые передаются только по материнской линии. Исследования мутаций в этих ДНК позволяют реконструировать историю биологического развития человечества, происхождение отдельных рас и народностей. Мутация может быть рецессивной, доминантной и полудоминантной в зависимости от состояния гена, в котором она произошла. Гены мутируют с определенной частотой, и ...
— концепция «2К» (кооперация и коммуникация) — эта концепция была разработана в 1900 г. русским ученым-философом П.А.Кропоткиным (1842—1921). В ней идет речь в основном о механизме биологической эволюции. . Термин «эволюция» (лат. evolutio — развертывание) в науке о живом представляет фундаментальное понятие для объяснения возникновения и развития всего живого. Эволюция подразумевает необратимое ...
... о противоречивом пути развития синтетической теории эволюции, о преодолении кризисов в научном познании, направленном на более полное и глубокое понимание закономерностей развития органического мира. Важно отметить, что основы научной теории эволюции заложил Ч. Дарвин. Как господствующее эволюционное учение дарвинизм существовал с 1859 до 1900 гг., т.е. до переоткрытия законов Г. Менделя. До ...
0 комментариев